
Abstract
Background Common genetic variants make a significant contribution to dimensional neurodevelopmental characteristics such as ADHD symptoms and cognitive abilities. The relevance and structure of these associations within a transdiagnostic sample referred for difficulties in cognition, attention and learning has not been demonstrated.
Methods Polygenic scores (PGS) derived from recent genome-wide association study (GWAS) data on ADHD and Intelligence were calculated from genome-wide genotyping data for 524 children and young people referred to the Centre for Attention, Learning and Memory (CALM). PGS-trait associations were tested via linear regression analyses, for a range of neurodevelopmental characteristics, and factor scores from a hierarchical model of psychopathology. Associations were explored with and without co-varying for socio-economic status (SES).
Results We found the expected associations within the sample between ADHD-PGS and ADHD symptom dimensions, and between Intelligence-PGS and IQ test scores. ADHD-PGS were also associated with broader externalising behaviours and with verbal intelligence, but the latter only if SES was not included as a covariate. Examining associations with hierarchical factor scores, we found a relationship between ADHD-PGS and a general mental health factor, externalising factor, and social maladjustment factor, however these associations did not survive co-variation for SES and correction for multiple comparisons. The Intelligence-PGS showed significant associations only with verbal and non-verbal cognitive skills, which were independent of SES; and no links were observed with ADHD traits or mental health factor scores.
Conclusion Our finding suggest that PGS derived from previous GWAS have research utility for paediatric transdiagnostic samples, showing both specific and general associations with neurodevelopmental traits. Future studies investigating PGS and cognitive and neural correlates within this cohort, as well as gene-by-environment interactions, may contribute to our understanding of developmental pathways and risk-resilience mechanisms in child mental health.
Key points
Common polygenic variation, quantified by PolyGenic Scores (PGS), contributes to dimensional neurodevelopmental characteristics. However, PGS relevance has not been demonstrated in a paediatric transdiagnostic sample.
We studied the associations between neurodevelopmental dimensions and ADHD PGS or Intelligence PGS, within a cohort of children referred for difficulties in cognition, attention and/or learning.
ADHD-PGS contributed to ADHD symptoms, as well as to externalizing behaviours, verbal intelligence, and hierarchical factors of mental health, with some of these associations being influenced by socioeconomic status (SES).
Intelligence-PGS in contrast, showed associations only with cognitive skills (verbal and non-verbal IQ), which were independent of SES; and did not link with ADHD traits or mental health factor scores.
Further research building on these findings will contribute to our understanding of developmental pathways and risk-resilience mechanisms in child mental health.
Introduction
Polygenic scores (PGS) are estimates of individual common genetic variation associated with a trait or disorder, and can provide insights into the genetic architecture and underlying mechanisms contributing to complex phenotypes (Andlauer & Nöthen, 2020; Choi et al., 2020). In particular, PGS have been extensively used to study the common polygenic contributions to a variety of neurodevelopmental traits in the general population, and neurodevelopmental diagnoses in case/control studies (de la Fuente et al., 2020; Ronald et al., 2021). Although traits and diagnoses are often investigated in isolation from each other, shared genetic contributions across disorders are well-recognised (Gidziela et al., 2023; Lee et al., 2019; Romero et al., 2022), indicating that PGS influence aspects of neurobiological and cognitive development relevant to a diverse set of behavioural and psychiatric vulnerabilities.
Neurodevelopmental difficulties often affect multiple domains of functioning that impact on learning, emotional-behavioural regulation, and social function, with complex within-sample profiles of strength and difficulty. Moreover, children with neurodevelopmental difficulties often do not meet criteria for a specific diagnosis, or meet criteria for multiple diagnoses. Recognising these complexities, those studying developmental difficulties and mental health have increasingly adopted a transdiagnostic framework (Astle et al., 2022). Previous studies of the associations between common genetic variation and specific neurodevelopmental traits or diagnoses are of unclear relevance to the large population of children with transdiagnostic difficulties. For example, case/control studies with strict inclusion/exclusion criteria may not be representative of the typical presentation of the trait of interest, if individuals with additional or subthreshold problems have been excluded. To close this gap, determining the associations between multiple PGS and a range of traits within samples of children who have heterogeneous and co-occurring difficulties may provide a more inclusive starting point for investigating links between genomic variation and neurodevelopmental characteristics.
With this framework in mind, the Centre for Attention Learning and Memory in Cambridge (CALM, http://calm.mrc-cbu.cam.ac.uk/) recruited a cohort of approximately 800 children recognised by educational or clinical professionals as having difficulties in attention, learning and/or memory (Holmes et al., 2019). This unique transdiagnostic paediatric sample provides an opportunity to study the factors (genetics and environmental) and mechanisms that drive diversity in neurodevelopment, independent of categorical diagnoses and their co-occurrence. The CALM cohort is a well-characterised sample from the point of view of behavioural features (Bryant et al., 2020; Holmes, Mareva, et al., 2021; Mareva et al., 2023; Williams et al., 2022), dimensions of cognitive and executive functions (Bathelt et al., 2018; Holmes, Guy, et al., 2021; Mareva & Holmes, 2019), dimensions of mental health (Guy et al., 2022; Holmes, Mareva, et al., 2021), and neuroimaging (Akarca et al., 2021; Siugzdaite et al., 2020). The current paper addresses the question of whether and how Attention-Deficit/Hyperactivity Disorder (ADHD) and Intelligence PGS are associated with relevant neurodevelopmental characteristics within the transdiagnostic CALM sample. We have focused on ADHD and Intelligence PGS because these traits reflect important aspects of behavioural and cognitive heterogeneity within the CALM cohort, and there are robust large-scale GWAS studies to draw on for planned analyses.
ADHD is a common neurodevelopmental disorder characterised by symptoms in either the inattentive or hyperactive/impulsive domains, or both (Posner et al., 2020). ADHD affects approximately 5% of children and 2.5% of adults, with a high heritability of approximately 70-80% estimated from twin-based studies (Faraone et al., 2005), and high comorbidity with other psychiatric conditions (Jensen & Steinhausen, 2015). The latest genome-wide association meta-analysis (meta-GWAS) of case/control studies has identified 27 genome-wide significant risk loci, including genes enriched for expression in early brain development, with 14 % of SNP heritability for ADHD (Demontis et al., 2023). Higher ADHD-PGS have been associated with ADHD diagnoses in independent case/control samples (Demontis et al., 2019), as well as with ADHD symptoms in population-based studies (Brikell et al., 2020; Demontis et al., 2023; Martin et al., 2014), suggesting that an ADHD diagnosis represents the extreme expression of one or more heritable quantitative traits. In addition, 84–98% of ADHD-risk variants are shared with other psychiatric disorders (Demontis et al., 2019, 2023).
In keeping with these pleiotropic associations, ADHD-PGS have recently been explored within a hierarchical, dimensional model of childhood psychopathology known as HiToP (Allegrini et al., 2020; Kotov et al., 2017; Waszczuk et al., 2020). The HiToP model proposes a p-factor of general psychopathology at the apex, under which more specific factors are described (Holmes, Mareva, et al., 2021; Michelini et al., 2019). The p factor is highly inheritable (50-60% from twin studies) and the model aligns well with current understanding of the higher order genetic structure of psychopathology. ADHD-PGS have consistently been shown to contribute to the p factor, as well as to sub-factors of externalizing behaviours and neurodevelopmental difficulties (Brikell et al., 2020; Pat et al., 2022; Riglin et al., 2020; Waszczuk et al., 2023).
A proportion of children with symptoms of ADHD or other behavioural difficulties also have cognitive and learning-related difficulties. The ability to think abstractly and problem solve independently of any previously acquired knowledge is known as fluid intelligence (Deary et al., 2021). Fluid intelligence is highly heritable, with a general factor of intelligence (g) accounting for about 58% of the genetic variance in cognitive traits (de la Fuente et al., 2020). A large-scale, meta-GWAS for the g factor of general intelligence has identified 205 genomic loci implicating brain-expressed genes involved in nervous system development, neuron differentiation, and synapse structure and activity (Savage et al., 2018). There is existing evidence that genomic variation contributing to intelligence also impacts on behavioural and psychiatric traits. Genetic correlation and Mendelian randomization analysis showed a negative correlation between intelligence-associated variants and ADHD and depression symptoms (Deary et al., 2021; Savage et al., 2018). In reverse, genomic variation quantified in association with behavioural traits (ADHD-PGS) has also been negatively correlated with cognitive abilities including general intelligence, attention, verbal and non-verbal reasoning, and working memory (Demontis et al., 2023).
In summary, there is existing evidence of convergent genetic contributions to cognitive (intelligence) and behavioural (ADHD) traits during childhood. However, it is not currently known whether, in the context of a transdiagnostic sample of children with neurodevelopmental difficulties, polygenic contributions are specific to a trait (e.g. ADHD-PGS associated with ADHD symptoms), or shared across traits (e.g. ADHD-PGS associated with additional behavioural dimensions, and aspects of cognitive function). The extent to which PGS show specific or shared associations within the CALM sample could provide insight into the inter-relationships between dimensions of neurodevelopmental difficulty at a behavioural or mechanistic level.
Thus, our first objective was to establish whether ADHD and Intelligence PGS showed associations with their respective “discovery” traits i.e. ADHD symptoms and cognitive test scores, within the CALM sample. Our second objective was to examine whether PGS associations were specific to these traits, or extended to additional behavioural traits (externalising and internalising problems) demonstrating cross-over between behavioural and cognitive dimensions. Thirdly, we explored whether PGS could contribute to our understanding of a hierarchical model of psychopathology (HiToP) within this population. All PGS-trait association analyses were conducted with and without co-varying for socioeconomic status (SES) to highlight associations which are maximally influenced by gene x environment interactions.
Materials and Methods
Participants
A cohort of 1000 children aged 5 to 18 years was recruited by the Centre for Attention, Learning and Memory (CALM, http://calm.mrc-cbu.cam.ac.uk/) between February 2014 and December 2021 (Holmes et al., 2019). The cohort consisted of 800 children referred by a health or educational professional because of difficulties related to attention, learning and/or memory, and 200 children not referred for having difficulties, recruited from the same schools as the referred children. This paper reports analysis of the referred children only. Comprehensive phenotyping was carried out for all children (for details see Holmes et al., 2019), and an optional DNA sample (saliva) was provided. A total of 775 samples were submitted for genotyping, from which 615 corresponded to the referred children. After QC filtering (see below), a final N=524 participants remained, with 361 males and 163 females, and average age of 114.1, SD 28.5, range 62-223 months (Table 1). Within this group of participants, 38% had received a diagnosis of any developmental or psychiatric disorder from community services, 22.5% had received a diagnosis of ADHD, 11.8% had dyslexia or were under speech and language therapy, and 7% had received a diagnosis for autism. Index of Multiple Deprivation (IMD; (The English Indices of Deprivation 2019, 2019) was used to approximate the socioeconomic status of the sample. Scores for postcode areas in the United Kingdom range from 1st to 32844th (most to least deprived).·The range of IMD for the sample indicated participants came from areas with varying degrees of deprivation, with an average ranking above the national median (Table 1). The decile distribution for IMD in this cohort is shown in Fig. S1.
Phenotypic measures
The measures used in the current analysis are listed in Table 2, together with descriptive data for each measure within the CALM PGS-analysed sample. For further details, consult the study protocol (Holmes et al., 2019). For details about the method for obtaining HiToP factor scores, see (Holmes, Mareva, et al., 2021). Briefly, higher-order dimensions of psychopathology within the CALM cohort were determined using Goldberg’s bass-ackward factor analytic method (Goldberg, 2006). This method is based on multivariate hierarchical models of 1 factor, 2 factors, 3 factors and so on, which enables the sequential, top-down extraction of dimensions whilst retaining the shared variance among the measures (the model ceases at the maximal number of factors that can be extracted). Three measures were included in this analysis: CPSF, SDQ and RCADS-P (Table 2). Because the RCADS questionnaire was introduced later into the CALM study protocol, HiToP factor analysis was conducted for the subgroup of participants with all measures available, for whom genetic data was available: N=218 individuals after QC filtering.
DNA extraction, genotyping, QC filtering, imputations, and PGS calculation
For detailed methods, see Supporting Material. In brief, DNA samples were collected from saliva using the Oragene® DNA self-collection kits (DNA Genotek Inc). DNA extraction was performed using prepIT·L2P Kit (DNA Genotek Inc) following manufacturer instructions. Genome-wide SNP genotyping was carried out (in 2 batches total N=775) using the Global Screening Array with shared custom-content (GSA, Illumina, Inc., USA) on an iScan instrument. The raw intensity data was subjected to genotype calling, extensive pre-imputation quality control and SNP imputation using an automated computational workflow described previously (Hong et al., 2020, 2021). A total of 18 samples were excluded: 7 because of low quality DNA or evidence of cross-contamination, 10 because genetic sex could not be determined unambiguously or in whom there was a discrepancy with the clinically reported sex, 1 because of excessive missing data (--mind 0.05). The LD pruned dataset was used for principal component analysis in PLINK (Purcell et al., 2007) along with the reference dataset of the 1000 Genome Project Consortium Phase 3 (The 1000 Genomes Project Consortium, 2015) to assign ethnic descent groups. “European descent” was assigned to 678 out of 757 samples across both genotyping batches; only these n=678 European-descent samples were used in the subsequent analyses. Genotype imputation was performed with MiniMac3 (Das et al., 2016) software using the “Haplotype Reference Consortium” v1.1 reference panel (McCarthy et al., 2016). Overall, this procedure resulted in 39,131,578 imputed SNPs.
We computed PGS for each individual as described previously (Choi et al., 2020). Allele status and effect-size estimates were taken from the summary statistics of the largest ADHD (Demontis et al., 2023) and Intelligence (Savage et al., 2018) meta-GWAS published to date. Variant QC filtering was performed on both GWAS databases: duplicate and ambiguous SNPs were removed, and only SNPs with MAF >0.05 and imputation quality r2 >0.8 were included. The target CALM sample did not overlap with the GWAS datasets. GWAS and CALM genotyping data were annotated on the GRCh37/hg19 genome build. VCF files from both genotyping batches were merged using BCFTOOLS. Non-referred participants’ data were removed at this point, prior to sample and variant filtering. 9 samples were removed due to extreme heterozygosity (more than 3 SD from the mean), and 24 closely-related individuals were also removed, giving a final sample size of N=524. Variants were filtered for duplicate SNPs, SNPs with MAF<0.01, P value for HWE < 1e-6, and genotyping efficiency <99%. Variant matching and recoding between the ADHD/Intelligence GWAS base and CALM target datasets was performed using R (version 3.5.3), resulting in 5,583,323 and 3,994,342 overlapping SNPs for ADHD and Intelligence, respectively. After SNP-clumping, PGS were computed for a variety of P-value thresholds (Pt) in the primary GWAS data (0.001, 0.05, 0.10, 0.20, 0.30, 0.40, 0.50) using PLINK.
Statistical analysis
PGS-trait association analyses were conducted in R (v3.5.3) for each Pt. ADHD and Intelligence PGS were treated as independent variables in linear regression models adjusting for sex, age, and the first 6 components of a PCA performed on the pruned SNP pool dataset to account for population stratification. Analyses were carried out with and without co-variation for index of multiple deprivation (IMD) because of the complex confounding links between PGS, socioeconomic status and behavioural dimensions. Significant PGS-trait associations were determined after Bonferroni corrections to account for multiple comparisons (Table S1). Sensitivity analyses including genotyping batch as a covariate in the regression analysis did not appreciably affect the PGS-phenotype associations.
The variance explained (r2) was derived from comparing results from the full model (including outcome phenotype and covariates) vs the null model (linear model with covariates only). P values that explained the highest phenotypic variance (max r2), with and without covariation with IMD, are shown in Table S2 and S3 for the behavioural and cognitive measures, in Table S4 and S5 for the HiToP factors, and in Table S6 for the ADHD diagnoses analysis. For comparison purposes, presentation of Results focuses on the Pt values that significantly explained the largest proportion of phenotypic variance for the greatest number of PGS-trait association studies: Pt= .4 (PGS-behavioural and cognitive measures), and Pt= .05 (PGS-HiToP factors).
Results
ADHD-PGS and Intelligence-PGS are associated with respective discovery dimensions within CALM
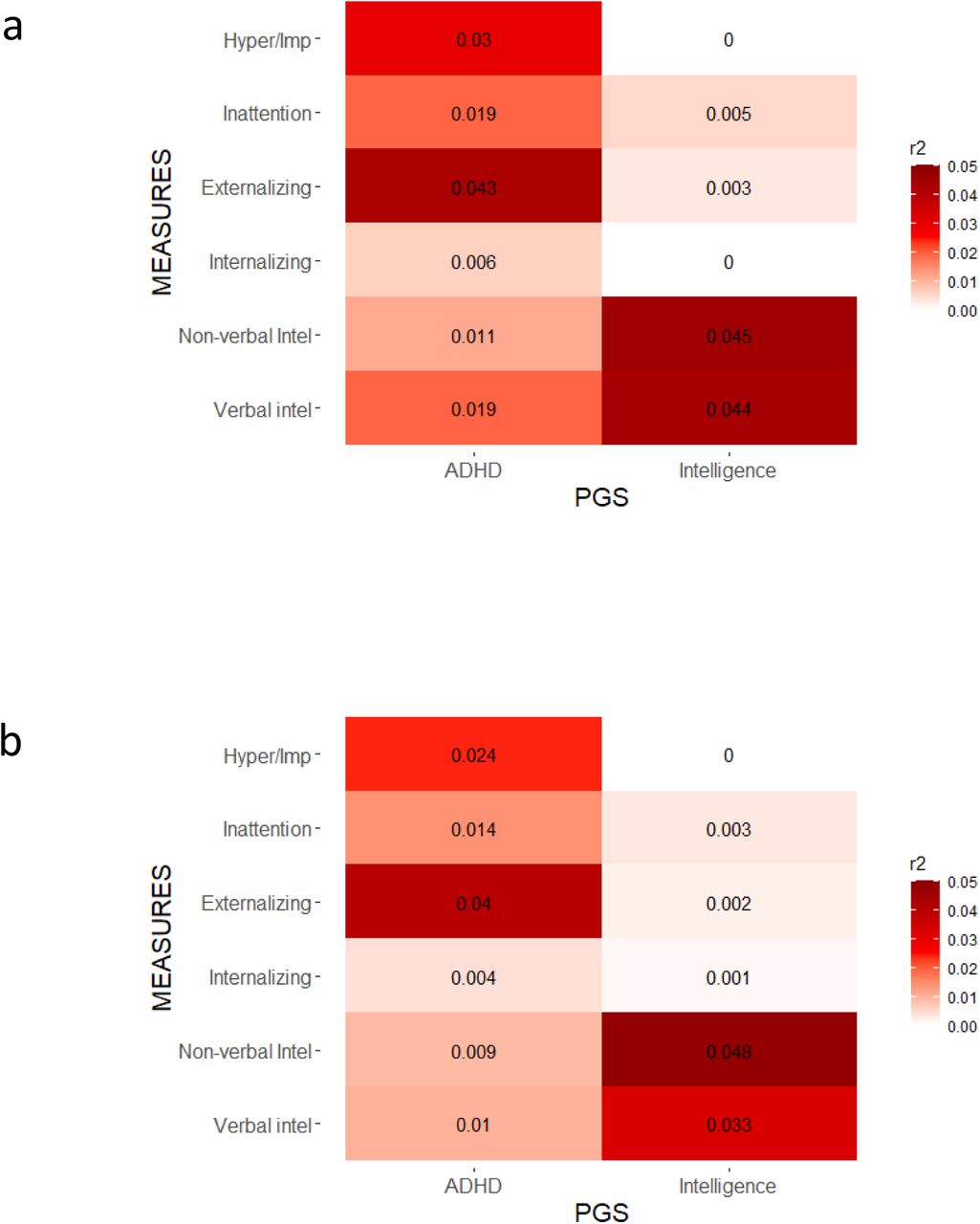
To assess the association between ADHD and Intelligence PGS with their respective behavioural and cognitive dimensions within the transdiagnostic CALM cohort, we conducted linear regressions with age, sex, and the first 6 ancestry PC as covariates. We present here results for Pt= .4 (see Materials and Methods for details), and considered significance after Bonferroni correction for multiple comparisons (Table S1). We found that ADHD-PGS were positively associated with Inattention (r2= .019; Beta=19114; p=1.44e-03) and Hyperactivity/Impulsivity scores (r2= .030; Beta=38672; p=3.06e-05) (Fig.1a,TableS2). Co-variation for IMD removed the significant association with Inattention (r2= .014; Beta=16060; p=9.91e-03), but retained the significant association with Hyperactivity/Impulsivity (r2= .024; Beta=35164; p=2.62e-04) (Fig 1b, Table S3). Co-variation for the presence/absence of ADHD diagnosis did not alter the relationships between ADHD-PGS and Hyperactivity/impulsivity or Inattention, including influence of IMD on the relationship with Inattention (Fig. 2, Table S6).

Heat map of r2 from linear regressions between ADHD and Intelligence-PGS and behavioural and cognitive dimensions within the CALM cohort, using a Pt=0.4 for PGS calculations. a) including sex, age and the first 6 ancestry PC (not including IMD) as covariates. b) including sex, age, the first 6 ancestry PC, and IMD as covariates. * p<.008, Bonferroni-corrected (Table S1). Hyper/Imp: Hyperactivity/Impulsivity, Intel: Intelligence.

Scatter plots showing the association between ADHD-PGS and a) Hyperactivity/Impulsivity and b) Inattention raw scores. ADHD diagnosis status is colour coded, with dark red being diagnosed for ADHD. A trendline was adjusted using linear model regression. The 95% CI is shaded in grey.
Intelligence-PGS were strongly positively associated with verbal (r2= .044; Beta=598764; p=7.95e-07) and non-verbal (r2= .045; Beta=199737; p=8.71e-07) intelligence test scores (Fig. 1a, Table S2). Co-variation for IMD did not affect the Intelligence-PGS associations (Fig 1b, Table S3).
ADHD-PGS associations extend beyond ADHD symptoms within CALM
To assess whether ADHD PGS contributed also to behavioural and cognitive dimensions beyond ADHD-related characteristics, we conduced linear regressions as described above. Again, we present here results for Pt= .4 using Bonferroni correction for multiple comparisons (Table S1). ADHD-PGS were strongly associated with externalising (r2= .043; Beta=34098; p=1.22e-06) but not internalising (r2= .006; Beta=14307; p=7.07e-02) difficulties, with and without co-variation for IMD (Figs 1a and 1b, Tables S2 and S3). ADHD-PGS were negatively associated with verbal intelligence (r2= .019; Beta=-161281; p=1.30e-03), but not with non-verbal intelligence (r2= .011; Beta=-40704; p=1.57e-02) (Fig 1a, Table S2). However, the association between ADHD-PGS and verbal intelligence was not significant after co-varying for IMD (Fig 1b, Table S3).
Intelligence-PGS were not significantly associated with ADHD symptom scores, externalising or internalising problems (Fig. 1, Tables S2 and S3).
ADHD-PGS contributes to hierarchical factors of mental health within CALM
To assess whether ADHD and Intelligence PGS were associated with mental health (considered as a tiered set of inter-related factors), we conducted regression analysis as described above, using the HiTOP factor scores calculated for a subgroup of participants within the CALM cohort (see Materials and Methods for details). We present here results for Pt= .05 using Bonferroni correction for multiple comparisons (Table S1). ADHD-PGS was positively associated with the level 1 general factor of psychopathology (r2= .028; Beta=2769; p=.014), the level 2 broad externalizing factor (r2= .029; Beta=2809; p=.010), and the level 3 social maladjustment factor (r2= .033; Beta=3051; p=.006) (Fig. 3a, Table S4). Covariation with IMD removed these significant associations: the association between ADHD-PGS and the general factor of psychopathology was no longer significant, whilst the associations between ADHD-PGS and broad externalizing and social maladjustment factors did not survive the Bonferroni correction (Fig 3b, Table S5).

{kind=link}
{kind=link}
{kind=link}
Heat map of r2 from linear regressions between ADHD and Intelligence-PGS and each factor of the three models of psychopathology within the CALM cohort, using a Pt=0.05 for PGS calculations. a) including sex, age and the first 6 ancestry PC (not including IMD) as covariates. b) including sex, age, the first 6 ancestry PC, and IMD as covariates. * p<.05, p<.025 and p<.017, respectively, Bonferroni-corrected (Table S1). Ext: externalizing, Int: internalizing, maladjust: maladjustment, Neurodevelop: neurodevelopmental factor.
Intelligence-PGS showed no significant association with any of the HiTOP psychopathology factors, with or without IMD as a covariate (Fig. 3a and b, Table S4 and S5).
Discussion
To the best of our knowledge, this is the first study of the polygenic contributions to neurodevelopmental characteristics within a paediatric transdiagnostic sample. Using the summary statistics of the largest meta-GWAS so far, we have derived PGS for ADHD and Intelligence for the transdiagnostic CALM cohort. We have examined the specificity and overlap of PGS associations with respective discovery phenotypes (ADHD symptoms, IQ test scores), additional behavioural characteristics (SDQ externalising and internalising scores), and factor scores derived from a hierarchical model of psychopathology (HiTOP). We have taken a first step toward exploring gene x environment interactions by assessing which PGS-trait associations are influenced by co-variation with socio-economic status (IMD).
First, we found that ADHD-PGS is associated with hyperactivity-impulsivity and inattention questionnaire scores, and Intelligence-PRS is associated with verbal and non-verbal intelligence test scores. This confirms that GWAS-derived metrics of common genomic variation are relevant to their expected dimensions of neurodevelopmental variation despite the inclusive ascertainment criteria, high levels of co-occurring diagnoses, and sub-diagnostic difficulties of the CALM cohort. Enrichment for multi-dimensional behavioural and cognitive difficulties within the cohort means that PGS-trait relationships can be observed despite a relatively small sample size. Although a high proportion of the cohort had received a diagnosis of ADHD from community services prior to recruitment to CALM, adding diagnostic status to this analysis did not alter results. These results provide a starting point for future explorations of the associations between PGS and cognitive and neuroimaging correlates, and longitudinal behavioural associations within the CALM cohort.
Second, we found that PGS derived from ADHD meta-GWAS statistics are associated with neurodevelopmental dimensions beyond the core ADHD diagnostic symptoms, extending into externalising difficulties and verbal intelligence. This is consistent with previous studies that have found that higher ADHD-PGS are also associated with elevated neurodevelopmental, externalizing, and depressive and anxiety symptoms in diagnostic (Stergiakouli et al., 2015) and population studies (Brikell et al., 2020; Du Rietz et al., 2018). There are a number of possible explanations for these results. The SDQ externalising score incorporates a hyperactivity subscale, so the association between ADHD-PGS and externalising scores may be driven by this subscale. The relationship between ADHD and language/verbal difficulties is well-documented both in CALM and in the broader literature (Hawkins et al., 2016; Korrel et al., 2017). A single dimension of behavioural difficulties that includes both hyperactivity and inattention, was strongly and negatively associated with pragmatic communication skills within the CALM cohort (Hawkins et al., 2016). The case groups contributing to ADHD-GWAS studies from which the ADHD-PGS is drawn likely included many individuals with ADHD plus additional behavioural symptoms plus reduced verbal intelligence, meaning that the ADHD-PGS summates genomic variation contributing to each of these separate dimensions (Demontis et al., 2023). The symptoms assessed by ADHD questionnaires (particularly for the inattention subscale) are diverse and incorporate cognitive and behavioural features likely to be influenced by cognitive abilities for at least some children, meaning cross-over associations of the ADHD-PGS with diverse neurodevelopmental dimensions. Lastly, genomic variation contributing to risk of ADHD symptoms may also contribute to risk of these additional phenotypes (a truly transdiagnostic genetic effect), evidenced by the significant genetic correlations between ADHD risk variants and IQ and educational attainment (Demontis et al., 2019). Future studies could test these alternative explanations.
In contrast, Intelligence-PGS showed a strong specific relationship with only verbal and non-verbal IQ test scores and did not cross-associate with any behavioural measures. This highlights the relative independence of general cognitive abilities and behavioural dimensions within this sample, and suggests separable aspects of genomic variation contribute to these broad domains. Previous analyses of cognition within the CALM cohort have identified three dimensions: phonological processing, processing speed and executive function (Holmes et al 2021) which can be explored with regard to PGS contributions in future analyses, accounting for developmental effects. PGS quantifies the load of polygenic variation but does not evaluate each individual’s unique combination of variants – application of newer techniques such as FUMA (Watanabe et al., 2017) and LAVA (Werme et al., 2022) may uncover important aspects of genomic diversity within the cohort, relevant to understanding different cognitive trajectories and their relationship with neurobiological mechanisms and learning/behavioural outcomes.
Whilst examining specific PGS-behavioural relationships has value and builds on previous literature, individual children within CALM (and children with co-occurring developmental difficulties in general) experience a combination of cognitive, behavioural, social and emotional challenges that may be better understood in an integrated fashion. The HiTOP model provides one such integrated framework, hence we have explored associations between HiTOP factors and PGS scores. Here again, we found that Intelligence-PGS had no predictive value in explaining psychopathological variation within the sample. ADHD-PGS analyses suggest that there are polygenic contributions to an overarching p factor within CALM, indexing general susceptibility to emotional-behavioural symptoms as has been shown previously (Brikell et al., 2020; Waszczuk et al., 2023). Interestingly, we did not detect any association between ADHD-PGS and the lower-level “neurodevelopmental” factor which encompasses inattention, hyperactivity/impulsivity and executive function. Instead, ADHD-PGS was associated with “social maladjustment”, encompassing conduct-related and peer-related problems assessed by the SDQ. We speculate that this observed association may be explained by the high comorbidity between ADHD and conduct disorders (Posner et al., 2020). A within-case analysis of individuals with comorbid ADHD and conduct disorders showed a positive linear relationship between ADHD-PGS and conduct problems, especially aggression (Hamshere et al., 2013). Alternatively, the relationship between ADHD-PGS and verbal abilities, rather than a direct effect on behavioural characteristics, may have also contributed to this observations. The importance of communication abilities (pragmatic skills in particular) and their relationship with behavioural symptomatology has been emphasised in previous CALM analyses (Hawkins et al., 2016), and future hypothesis-driven studies could investigate the links between ADHD-PGS and language development. However, it is important to note that the sample available for PGS-HiTop analysis was likely under-powered, and the specificity of associations should thus be treated with caution.
Within the CALM sample, socio-economic status (approximated via the Index of Multiple Deprivation) is an important demographic factor which is known to contribute to behavioural symptom likelihood and also to be correlated with polygenic variation (Abdellaoui & Verweij, 2021). Hence it is possible that observed PGS-behavioural associations are confounded by the influence of SES. To explore this, we conducted association analyses with and without IMD co-variation. We found that the Intelligence-PGS associations with verbal and non-verbal intelligence scores were insensitive to IMD, suggesting that there is relatively little environmental influence on general aspects of cognition (Rask-Andersen et al., 2021). Associations between ADHD-PGS and inattention, verbal intelligence and HiTop psychopathology factors were all weakened (or rendered non-significant) when co-varying for IMD. This may indicate that the observed associations are artefacts of co-variation, or that the sample is under-powered to detect significant associations once co-variates are accounted for, or alternatively that there are important gene x environment interactions at play. The association between ADHD-PGS and hyperactivity is not sensitive to IMD covariation, supporting a more direct and robust relationship between genomic variation and this symptom dimension, consistent with previous studies which have also found ADHD-PGS to be associated with hyperactivity/impulsivity more consistently than inattention (Brikell et al., 2020). Future studies could explore gene x environment interactions within this sample by assessing the impact of environmental risk factors for ADHD, such as parents’ education and prenatal/perinatal risk factors, on the emotional-behavioural and cognitive measures (Leffa et al., 2023).
In summary, we have obtained relevant PGS for a unique transdiagnostic sample, and found that they contribute significantly and specifically to behavioural characteristics and cognitive abilities. This is a first step toward investigating links between genomic variation, brain development and cognitive processes contributing to learning and behavioural problems within this population.
Data Availability
Behavioural and cognitive data is available online through the CALM Managed Open Access repository: https://portal.ide-cam.org.uk/overview/1158. Genetic data is available via collaboration with CALM.
Data Access
Behavioural and cognitive data is available online through the CALM Managed Open Access repository: https://portal.ide-cam.org.uk/overview/1158. Genetic data is available via collaboration with CALM.
Ethics
All procedures complied with the ethical standards of the national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008. All procedures involving human participants were approved by the National Health Service, Health Research Authority NRES Committee East of England (REC: 13/EE/0157, IRAS 127675).
Acknowledgments
D. Astle and K. Baker are supported by the Medical Research Council (G116768). A. Santangelo was supported by the Isaac Newton Trust / Wellcome Trust ISSF / University of Cambridge Joint Research Grants scheme. All research at the Department of Psychiatry in the University of Cambridge is supported by the National Institute for Health and Care Research Cambridge Biomedical Research Centre (NIHR203312) and the NIHR Applied Research Collaboration East of England. Genotyping was carried out within the “Healthy minds 0-100 years: Optimising the use of European brain imaging cohorts (Lifebrain)” project, funded by EU Horizon2020 (Grant agreement number: 732592). We are grateful to Professor Sadaf Farooqi and her laboratory for DNA extractions and storage, and to NIHR BRC-MRC BioRepository for sample archiving.
The Centre for Attention Learning and Memory (CALM) research clinic is based at the Medical Research Council (MRC) Cognition and Brain Sciences Unit, University of Cambridge, funded by the MRC through UKRI. The lead investigators are Duncan Astle, Kate Baker, Susan Gathercole, Joni Holmes, Rogier Kievit and Tom Manly. Data collection is assisted by a team of researchers and PhD students that includes Danyal Akarca, Joe Bathelt, Marc Bennett, Madalena Bettencourt, Giacomo Bignardi, Sarah Bishop, Erica Bottacin, Lara Bridge, Diandra Brkic, Annie Bryant, Sally Butterfield, Elizabeth Byrne, Gemma Crickmore, Edwin Dalmaijer, Fánchea Daly, Tina Emery, Laura Forde, Grace Franckel, Delia Furhmann, Andrew Gadie, Sara Gharooni, Jacalyn Guy, Erin Hawkins, Rebeca Ianov-Vitanov, Christian Iordanov, Agnieszka Jaroslawska, Sara Joeghan, Amy Johnson, Jonathan Jones, Silvana Mareva, Jessica Martin, Elise Ng-Cordell, Sinead O’Brien, Cliodhna O’Leary, Joseph Rennie, Andrea Santangelo, Ivan Simpson-Kent, Roma Siugzdaite, Tess Smith, Stepheni Uh, Maria Vedechkina, Francesca Woolgar, Natalia Zdorovtsova, Mengya Zhang. The authors wish to thank the many professionals working in children’s services in the South-East and East of England for their support, and to the children and their families for giving up their time to visit the clinic.
References
{kind=link}
{kind=link}



