
Abstract
In 2015, Brazil reported an outbreak identified as Zika virus (ZIKV) infection associated with congenital abnormalities. To date, a total of 86 countries and territories have described evidence of Zika infection and recently the appearance of the African ZIKV lineage in Brazil highlights the risk of a new epidemic. The spectrum of ZIKV infection-induced alterations at both cellular and molecular levels is not completely elucidated. Here, we present for the first time the gene expression responses associated with prenatal ZIKV infection from ocular cells. We applied a recently developed non-invasive method (impression cytology) which use eye cells as a model for ZIKV studies. The ocular profiling revealed significant differences between exposed and control groups, as well as a different pattern in ocular transcripts from Congenital Zika Syndrome (CZS) compared to ZIKV-exposed but asymptomatic infants. Our data showed pathways related to mismatch repair, cancer, and PI3K/AKT/mTOR signaling and genes probably causative or protective in the modulation of ZIKV infection. Ocular cells revealed the effects of ZIKV infection on primordial neuronal cell genes, evidenced by changes in genes associated with embryonic cells. The changes in gene expression support an association with the gestational period of the infection and provide evidence for the resulting clinical and ophthalmological pathologies. Additionally, the findings of cell death- and cancer-associated deregulated genes raise concerns about the early onset of other potential pathologies including the need for tumor surveillance. Our results thus provide direct evidence that infants exposed prenatally to the Zika virus, not only with CZS but also without clinical signs (asymptomatic) express cellular and molecular changes with potential clinical implications.
Introduction
In 2015, Brazil reported an outbreak identified as Zika virus (ZIKV) infection, transmitted mainly by Aedes mosquitoes1,2. The transmission of this disease, associated with Guillain-Barré syndrome and microcephaly, has been registered in different regions of the world. To date, a total of 86 countries and territories have described evidence of Zika infection (transmitted by mosquitoes) 1 and, more recently -in 2019, the appearance of the African ZIKV lineage in Brazil highlight the risk of a new epidemic3.
Congenital Zika Syndrome (CZS) was identified due to the increased incidence of congenital abnormalities associated with ZIKV infection, which include morphological, behavioral, neurological4,5,6,7 and ocular impairments8,9,10,11,12,13. Notably, not only ZIKV antigens have been detected in ocular tissue samples14, but also ZIKV has the potential to survive for long periods in ocular tissue and it is detectable in ocular fluids (e.g. lacrimal fluid)15,16,17.
Ocular lesions may result from defects of eye development that may have occurred at different stages of embryogenesis8,9,10,11,12,13,14. On the other hand, the eye has been described as a specialized CNS compartment, since the optic nerve and retina extend from the diencephalon during embryonic development, being able to display symptoms of neurodegenerative diseases18.
ZIKV infection might yield outcomes that will only be manifested later in life19, requiring long-term monitoring of all infants prenatally exposed to ZIKV20,21. Considering the need for after birth surveillance of infants with possible ZIKV congenital exposure, we have standardized an impression cytology methodology and study protocol for analysis of molecular alterations resulting from ZIKV infection in ocular tissue22.
Here we present the first study of global gene expression responses of CZS from ocular samples captured by impression cytology. The ocular profiling revealed significant differences between exposed and controls, as well as a different pattern in ocular transcripts from CZS compared to asymptomatic infants. Congenital ZIKV exposure triggered the regulation of pathways related to mismatch repair, cancer, and PI3K/AKT/mTOR signaling and genes probably causative or protective in the modulation of ZIKV exposure. Altogether, our results enlighten the understanding on the molecular effects of ZIKV exposure during human embryonic development, getting attention to the need for surveillance of all exposed children, even when born without clinical signs.
Material and Methods
Ethical statement and clinical aspects
This study was carried out in accordance with the ethical principles taken from the Declaration of Helsinki and written informed consent was obtained from the parents of babies under Institutional Review Board and Ethics Committee of Universidade Federal Fluminense approved protocols (approved protocols register number - CAAE: 79890517.6.0000.5243). Ocular surface samples were obtained from a total of twelve infants diagnosed according to the CZS diagnostic criteria referred to the Pediatric Service of the Hospital Universitário Antonio Pedro, Universidade Federal Fluminense (HUAP-UFF), Brazil. All children have been followed by periodical clinical examinations including neurological and ophthalmological screening. From eight babies exposed to the Zika virus during pregnancy, four presented clinical characteristics corresponding to CZS while four did not present clinical signs at the time of ocular cell collection (asymptomatic infants). Control samples were obtained from four unaffected infants, with no clinical signs and no exposure to ZIKV during gestation.
Impression cytology and ocular surface cells capture
Ocular surface samples from the infants exposed to ZIKV were obtained through optimized and non-invasive impression cytology method using a hydrophilic nitrocellulose membrane as previously described22. Additionally, cytospin preparations were done to enable concentration of single cells in suspension on a microscope slide and morphological evaluation (qualitative and quantitative) of potential cell changes. All procedures were described in detail in our previous report22.
RNAseq and Data analysis
Cell suspensions (9 μL/sample) containing a maximum number of 150 single cells/sample (nuclei) were placed in individual 0.1 mL PCR tubes with 2.5 μl 1 x PBS (excluding calcium and magnesium) and cells were processed through the Single Cell RNAseq Service (SingulOmics/ Novogene). Processing included RNA isolation, cDNA synthesis and amplification, library preparation, and sequencing (10 million paired-end reads per sample)22. Reads from RNA-seq were subjected to quality trimming using Trim Galore (version 0.5.0) and aligned to human reference genome (GRCh38.93) using STAR (version 2.6.1a; options: --outSAMattrIHstart 0 --outSAMstrandField intronMotif --outFilterIntronMotifs RemoveNoncanonical --alignIntronMin 20 --alignIntronMax 1000000 --outFilterMultimapNmax 1)23. Duplicated reads were discovered using Picard tools (version 2.0.1) and removed. Gene annotations (gtf file; version GRCh38.93) were obtained from Ensembl. FPKM values of genes were estimated using cufflinks (version 2.2.1)24. Counts of genes were estimated using Ht-Seq (v0.10.0)25 and Ensembl gene annotation file (gtf file; v. GRCh38.93). Moreover, genes with less than 5 normalized counts in one or more samples are excluded from the analysis. The statistical analyses were performed in R v 3.5.0. We processed the data using DESeq2 v1.14.126. The reads counts were normalized, the fold change between groups was calculated, and the differentially expressed genes were selected by adjusting |Log2 FC|> 1 and False Discovery rate (FDR) corrected p-value (Benjamini–Hochberg) <0.05. We then analyzed a set of genes (top 5%-fold change) to find enriched Gene Ontology (GO) terms with respect to the complete set of tested genes (8583). We used the Gorilla web service27 with the running mode “Two unranked lists of genes” and default P-value threshold (1⍰×⍰10−3).
Quantitative Analysis of transcriptomic Data and Statistical Analysis
Data were processed using Perseus software (v1.4.1.3, Max Planck Institute of Biochemistry, Martinsried, Germany)28. Principal component analysis (PCA) was performed as exploratory data analysis approach in all quantified transcripts. One-way analysis of variance (ANOVA) was used for multiple testing while t-tests were performed for the comparison between two groups. For both tests, p-value< 0.05 were considered statistically significant. Regression and correlation analyses were also performed for the obtained results. The correlations between variables were defined by the Pearson (Perseus) coefficients, and p values less than 0.05 were considered significant. Protein-protein interaction networks were constructed using STRING29 software with High confidence parameter (0.700). Pathway and disease enrichment analysis were also carried out ToppFun function of ToppGene30 suite with FDR correction of 1%.
Results
Clinical aspects
Samples were collected from both eyes of infants (eight boys and four girls) with median age of 21 months (Supplemental Table 1). Four infants were ZIKV exposed and presented CZS according to the CZS diagnostic criteria (ZIKV/CZS: four infants with positive PCR for Zika virus in gestation and presence of clinical signs which included ocular abnormalities and microcephaly – ZIKV infection predominantly in the first trimester), four infants were ZIKV exposed (positive PCR for Zika virus during gestation; maternal ZIKV infection occurring in the second and third trimester) but no clinical signs identified at sample collection time (ZIKV asymptomatic infants) and four unaffected and ZIKV unexposed infants (control samples - CTRL with negative PCR and without clinical signs; Figure 1).

A. Schematic figure representing the different groups studied - Exposed (CZS and ZIKV) and not exposed (control) prenatally to the Zika virus. B. Cell collection procedure and experimental design.
Ocular Cell surface abnormalities
The morphology of cells collected from exposed and non-exposed infants was evaluated in parallel studies22,31. Changes associated with cell degeneration and death were clearly detected in cells from ZIKV-exposed patients, predominantly in CZS, compared to controls and included different levels of keratinization, pyknosis, karyolysis, anucleation and cytoplasmic vacuolization (Figure 2). Quantification of these cytoplasmic and nuclear alterations demonstrated a significant higher alteration in infected infants (Figure 2).

(A, B) Representative cells from uninfected group show normal morphology. (C-I) Nuclear and cytoplasmic alterations observed in cells collected from ZIKV (infected with no clinical signs) or ZIKV/CZV (diagnosed congenital Zika syndrome) children. Morphological changes were characterized by pyknosis (C, E, H), keratinization (C, D, E), anucleation (D, G), karyolysis (F), vacuolization (C, H) and cell fragmentation (H). Quantitative analyses (I, J) show increased number of conjunctival cells with morphological changes. Cells were collected by impression cytology, cytocentrifuged and stained with Diff-Quik or toluidine blue. For each group, 100 cells were analyzed in 2 different slides and scored for morphological changes. Data represent mean ± SEM. * P < 0.001.
RNAseq profiling
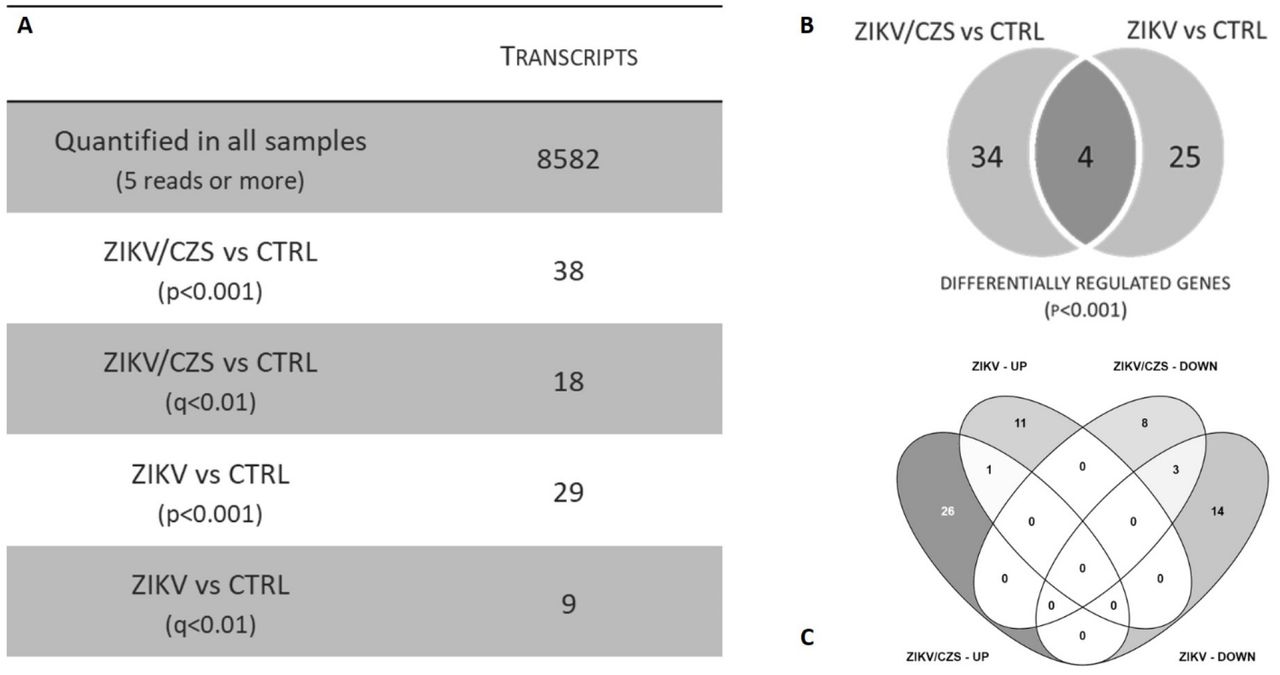
Transcriptome analysis showed 8582 transcripts quantified in all samples (> 5 reads in all samples; Figure 3A). Principle Component Analysis of these transcripts in all conditions with a non-supervised distribution showed no apparent difference among the samples.

A. Descriptive analysis table of transcripts quantified and regulated. B. Venn diagram of DRG (p<0.001) according exposed group. C. Differentially expressed gene per exposed group by VENNY analysis.
Ocular cells from exposed infants, ZIKV/CZS and ZIKV, showed 63 differentially expressed genes [p-value<0.001; 27 differentially expressed genes if considering adjusted p-value <0.01], when compared to the unaffected group (CTRL) group. However, the ZIKV exposed groups are heterogeneous within the genes differentially expressed between the two of them. Thus, we proceeded to contrast each of the exposed groups with the unexposed controls (Figure 3C).
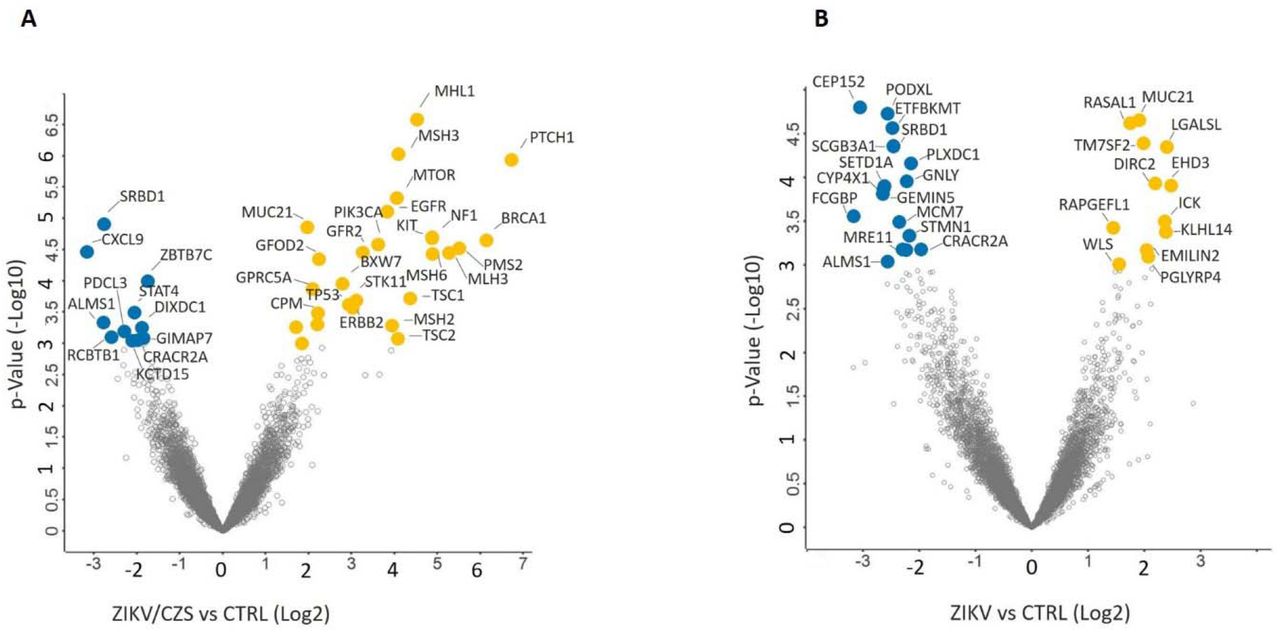
A total of 38 genes are differentially expressed in ocular cells from ZIKV/CZS compared to CTRL (Figure 4A and Table 1) while 29 genes are regulated in ZIKV compared to CTRL (Figure 4B and Table 2). Only 4 genes (SRBD1, MUC21, ALMS1 and CRACR2A; p<0.001) are differentially regulated in ocular cells from both exposed groups when compared to CTRL; Figure 3B).

Ocular transcripts differentially expressed comparative analysis A. CZS sample compared to CTRL samples. B. ZIKV without clinical signs sample compared to CTRL samples. Volcano plot showed transcripts up-regulated (yellow) and downregulated (blue).
Comparative transcriptomics
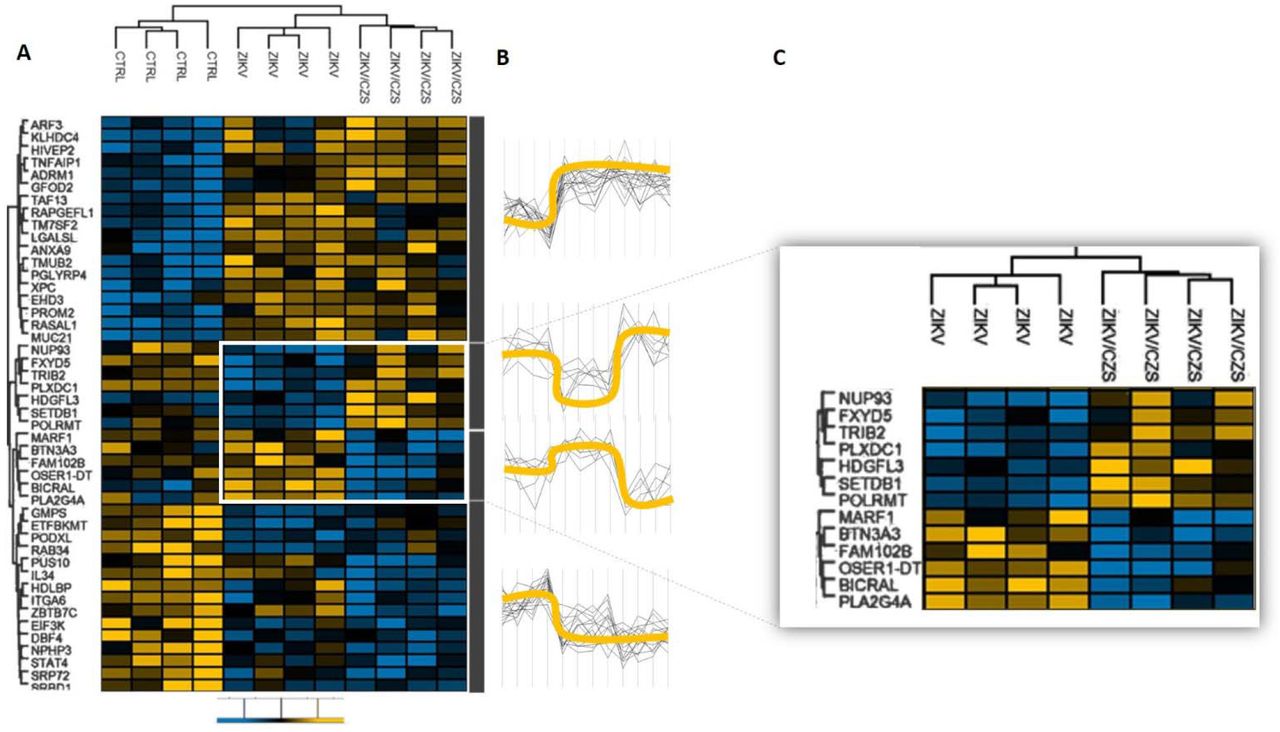
Comparison among ZIKV/CZS, ZIKV and CTRL groups revealed four well-defined hierarchical clusters (Figure 5). The first cluster showed a clear association of genes ARF3, KLHDC4, HIVEP2, TNFAIP1, ADRM1, GFOD2, TAF13, RAPGEFL1, TM7SF2, LGALSL, ANXA9, TMUB2, PGLYRP4, XPC, EHD3, PROM2, RASAL1 and MUC21 in most cases downregulated in CTRL infants and upregulated in all ZIKV exposed infants (either CZS and ZIKV, without clinical signs). The second cluster showed association of genes NUP93, FXYD5, TRIB2, PLXDC1, HDGFL3, SETDB1 and POLRMT in most cases downregulated in CTRL and ZIKV/CZS and upregulated in ZIKV exposed infants. The third cluster showed association of genes MARF1, BTN3A3, FAM102B, OSER1-DT, BICRAL and PLA2G4A in most cases downregulated in ZIKV/CZS and upregulated in CTRL and ZIKV exposed infants. The fourth-cluster showed association of genes GMPS, ETFBKMT, PODXL, RAB34, PUS10, IL34, HDLBP, ITGA6, ZBTB7C, EIF3K, DBF4, NPHP3, STAT4, SRP72 and SRBD1 in most cases downregulated in all ZIKV exposed infants (either CZS and ZIKV, without clinical signs) and upregulated in CTRL infants.

A. Changes in expression levels are displayed from blue (downregulated) to yellow (upregulated). B. Expression levels between groups represented by yellow line. C. Highlighted are the differential gene expression between the ZIKV groups exposed. Heat map were generated, and hierarchical clustering was performed using Perseus based on log2 fold-change data. Scale bar = Z score.
Functional enrichments
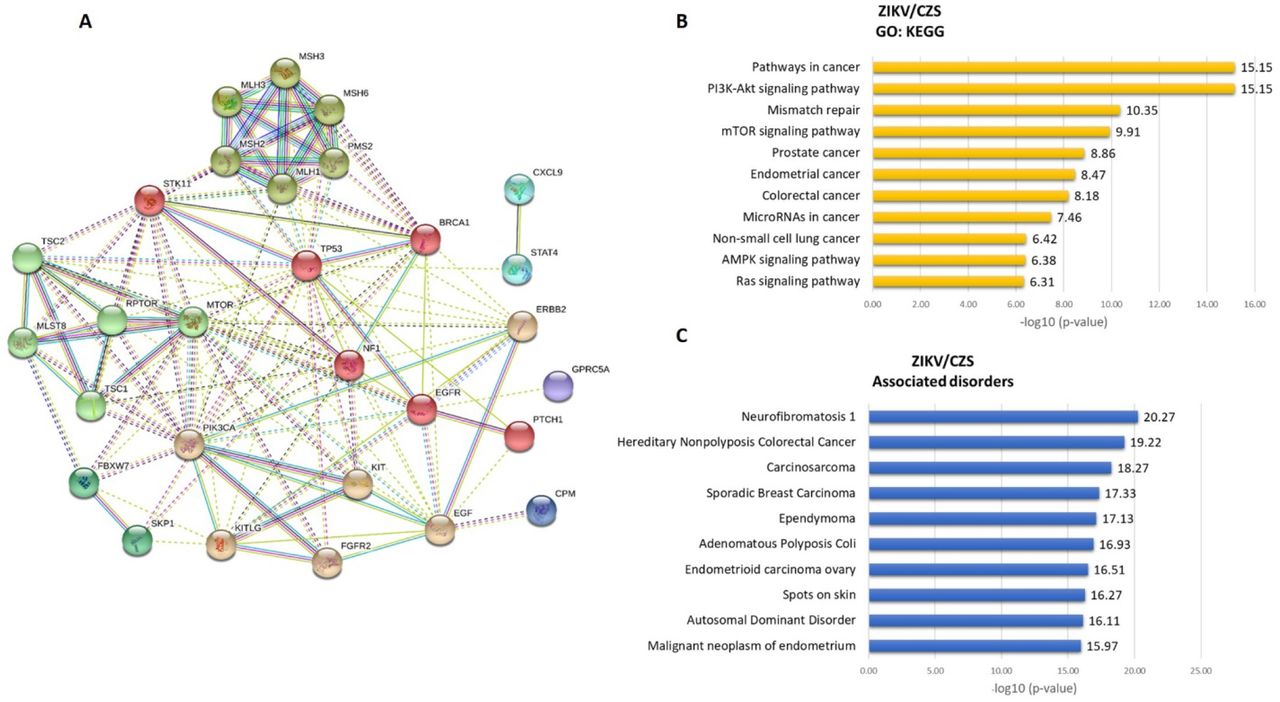
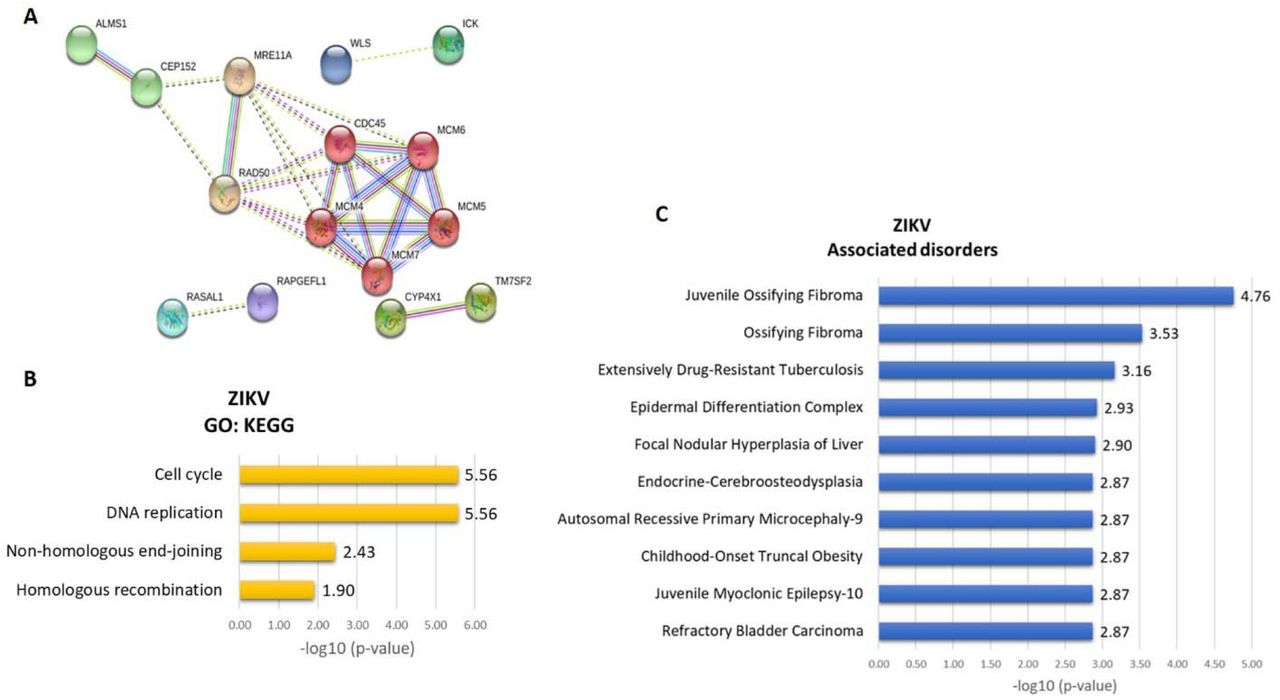
We found a significant enrichment of Gene Ontology (GO) annotations among the 63 differentially expressed transcripts detected in the RNA-Seq experiment (Figures 6 and 8). The top three most significantly enriched GO differentially regulated transcripts in ZIKV/CZS compared to CTRL (Supplemental Table 2) are “PI3K-Akt signaling pathway”, “Pathways in cancer”, “Mismatch repair” and “mTOR signaling pathway” (Figure 6). As the most significantly enriched GO from ZIKV compared to CTRL group are “DNA replication”, “Cell cycle” and “Non-homologous end-joining” (Figure 8; Supplemental Table 3).

A. The thickness of the lines connecting the circles indicates the number of interactions between genes included in each Gene Ontology (GO) sub-category represented by each circle. B. Top 10 enriched GO interaction network according STRING/KEGG analysis. C. Top 10 associated disorders according Toppgene analysis.

A. Top 10 embryogenesis stages related. Top 10 neurological process associated. C. Top 10 ocular phenotype associated.

A. Deregulated genes in ZIKV babies. The thickness of the lines connecting the circles indicates the number of interactions between genes included in each Gene Ontology (GO) sub-category represented by each circle. B. Top 10 enriched GO interaction network according STRING/KEGG analysis. C. Top 10 associated disorders according Toppgene analysis.
Discussion
The eye as neuroepithelial model to study Zika virus infection
Human sensory organs and cranial sensory ganglia are functionally diverse but share a common developmental origin from neural progenitor cells. During the different stages of embryogenesis - through a process that requires a rigorous integration between multiple signaling pathways, repressive transcription factors, cell target specification and morphogenesis - the inner ear, olfactory epithelium, lens, and sensory neurons are gradually originated32. Besides, the eye – a specialized CNS compartment - has been proposed as a model for studies in neurodegenerative diseases18,22. We have recently described a novel non-invasive method that enables collecting ocular cells from children22 as alternative neuroepithelial model to study Zika virus infection and here we applied this technique, for the first time, to investigate the extent of gene expression dysregulation in the ocular tissue of zika virus prenatally exposed children with and without CZS.
Differentially expressed genes involved in neurogenesis, eye development, and neurological disorders
Molecular and cellular events leading to abnormal nervous system development in CZS infants are poorly understood, but the host’s response to ZIKV insults is likely to be modulated by the combined effects of multiple genes. Several genes emerged here as differentially expressed in both groups of exposed infants (CZS and ZIKV), many of which are involved in neurogenesis and ocular development (including mRNA expression in embryonic tissues and primordial stem cells). Indeed, the impact of ZIKV infection on neuronal regulation has already been shown experimentally33,34,35, 36, 36,37 and is consistent with the hypothesis that changes in central nervous system embryogenesis that predict adverse outcome as observed in CZS are modulated by deregulation in several signaling pathways (Figures 6 and 7; Supplemental Tables 4, 5 and 6). Here we describe several genes and their biological contexts associated to neurodevelopment (gene details38 in Boxes 1A, 1B and 2A) and highlight a likely implication of a deregulation of these molecules in CZS (Figures 6 and 7).
1A. Upregulated genes description. 1B. Downregulated genes description.
2A. CZS associated genes. 2B. Asymptomatic ZIKV associated genes.
Collectively, the identity and function of these genes support the hypothesis that Zika virus infection in the first trimester of pregnancy affects precursor cells with effects that extend during the fetal developmental period and can still be seen in the differentiated stages of the cells in infants after birth (Figure 9). Ocular surface cells collected around 21 months after birth contain changes in embryonic marker genes and may be a representation of the interaction of precursor neuronal cells with the mechanisms of Zika virus infection during embryogenesis. Surprisingly, however, we also observed gene expression dysregulation of neurological genes in ZIKV groups associated to several neurological syndromes, developmental delay, intellectual disability, autism spectrum disorders, schizophrenia, and/or ocular impairment that we hypothesize might be relevant in these children. As demonstrated by several studies, there has been an advance in knowledge about the pathophysiology of ZIKV infection in the embryonic development of the CNS33, 39, 34, 35, 36, 40, 37. However, the knowledge about which pathways are associated with the pathogenic mechanism remains poorly understood. This directly interferes in the application of a more effective therapeutic approach, considering the serious clinical consequences for ZIKV exposed individuals41, and evidenced here for prenatally ZIKV exposed - both asymptomatic and CZS patients.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dysregulation of genes associated to Congenital Zika Syndrome
Comparative transcriptomics into ZIKV exposed infants and controls showed a downregulation of six genes also associated to CZS phenotypes (Figure 5). Here, we hypothesize that haploinsufficiency of these genes may have contributed to the CZS phenotype. Clinical variants of the OSER1-DT gene are associated with developmental delay/intellectual disability, autism spectrum disorders and congenital anomalies42; the BTN3A3 gene is involved in immunity, cell proliferation and development43; the FAM102B gene possesses in vivo function unknown38 but is highly expressed in retina44 and associated to glaucoma45; the PLA2G4A gene is involved in inflammatory responses and all-trans-retinoic acid signaling in brain and moreover, transcripts are in embryonic tissues, blood brain barrier, extraembryonic mesoderm46; the MARF1 gene required to protect from DNA double-strand breaks38 and the BICRAL gene, a component of chromatin remodeling47 (See more gene details38 in Box 2A).
Genes related to response to infection mechanisms
We identified some deregulated genes in ZIKV exposed infants (either CZS or asymptomatic) associated to immune response mechanisms. In first cluster, for example, the upregulated HIVEP2 gene possess enhancer elements of numerous viral promoters (such as those of SV40, CMV, or HIV1) and ANXA9 gene - which expression is regulated by IL-22 in keratinocytes48,49. On the other hand, genes shown in fourth cluster are downregulated, such as IL34 gene (related to innate immunity and in inflammatory processes50; EIF3K gene (overexpressed in T-lymphocyte) and STAT4 gene (See more gene details38 in Boxes 1 and 2). Several studies showed to the association of these genes with virus infection, such as papilloma virus 16 (HPV16), HIV infection and life cycle, Maraba, Hepatitis C and West Nile virus38.
PI3/AKT/mTOR signaling in CZS
We found upregulation of several members of the PI3/AKT/mTOR signaling pathway (BRCA1, EGF, EGFR, FGFR2, KIT, KITLG, MLST8, MTOR, PIK3CA, RPTOR, STK11, TP53, TSC1 and TSC2 genes) in ocular samples from CZS infants. PI3K-AKT signaling pathway has an important role in brain growth and it is the cause of many brain overgrowth disorders51. The PI3K / AKT pathway is crucial for normal neural stem cells in the brain, balancing the maintenance of their multipotent, renewing and proliferating rather than differentiating and becoming inactive52,53,54. Subsequent to an injury, neural stem cells enter a repair phase and express high PI3K levels to increase proliferation55. A critical protagonism for the CDK1-PDK1-PI3K/Akt kinase signaling pathway has been suggested in the regulation of self-renewal, differentiation, and somatic reprogramming55,56. As observed during mouse embryo development, AKT activity has a role in mitosis and myogenic and neural differentiation and/or survival57. The causes of microcephaly by ZIKV are not fully understood but recently, it is reported that ZIKV protein NS4A and NS4B inhibit Akt-mTOR signaling pathways, resulting in upregulation of autophagy, and promoting viral replication58. It is been showed that ZIKV infection of human fetal neural stem cells cause inhibition of the Akt-mTOR pathway, leading to defective neurogenesis and aberrant activation of autophagy58. ZIKV proteins to improve viral replication by blocking two innate pathways: interferon (IFN) and mTOR signaling (that normally would inhibit viral replication). The NS5 protein promotes degradation of the interferon effector STAT2, which would then promote transcription of interferon pathway genes. NS4A and NS4B inhibit mTOR signaling emanating from receptor tyrosine kinases. These effects of ZIKV proteins also to inhibit neurogenesis, endorsing cell death59.
Dysregulation of DNA Repair and cellular death
We found upregulated Mismatch Repair (MMR) pathway (MLH1, MLH3, MSH2, MSH3, MSH6 and PMS2 genes) in ocular samples from CZS infants. It has been suggesting that DNA damage can cause premature differentiation and apoptosis60. For instance, the regulation of Msh2 protein levels in the ESC interferes the decision of engaging on DNA repair pathways or undergoing apoptosis61. High levels of Msh2 expression - as detected in CZS infants in our pilot study - may lead to apoptotic responses62. Here, we propose that genotoxic stress and DNA repair responses influence neurodegeneration in the CZS, considering the that poorly repaired DNA damage can be a trigger for apoptosis, and the consequent loss of neurons by apoptosis can result in neurodegeneration, and moreover persistent DNA damage can result in defective neurogenesis, such as observed in CZS. To date, no association has been demonstrated between MMR genes and Zika virus infection in patients. However, this relationship has already been shown experimentally39 and others studies on the role of MMR in viral infections. Levels of MSH2 and MLH1 mRNA were dramatically increased in cells after avian leukosis virus infection, suggest that viral infection may promote the increased expression of MSH2 and MLH1 genes61. Post-replicative MMR factors have been shown to be recruited into Epstein-Barr virus (EBV) replication compartments, repairing errors that arise during viral replication or inhibiting recombination between conflict sequences. EBV DNA polymerase has an intrinsic 3’ to 5’ exonuclease activity and the fidelity of viral genome replication is believed to be high, therefore the MMR system might increase the integrity of the viral genome rather than to activate cytotoxic responses63. For effective HSV-1 replication in normal human cells, MSH2 and MLH1 are equally required while MLH1 knockdown inhibits early viral gene expression and MSH2 seems to act at a later stage in the viral life cycle64. In fact, morphological changes associated with cell degeneration and death were observed in the cells of infants exposed to ZIKV, both in asymptomatic and in CZS, being predominant in this group. These results corroborate our molecular results and highlight the importance of further studies.
Dysregulation in DNA replication, cell cycle and Non-homologous end-joining recombination
We identified downregulation of Minichromosome Maintenance protein complex (MCM) in asymptomatic ZIKV infants. MCM is a DNA helicase essential for genomic DNA replication that consists of six gene products, MCM2–7, which form a heterohexamer65. As a critical protein for cell division, MCM is also the target of various checkpoint pathways and deregulation of MCM function has been linked to genomic instability and a variety of carcinomas65. In zebrafish experimental model has demonstrated that mcm5 is down regulated in differentiated cells but is preserved in retinal stem cells regions. A regular depletion of maternally derived protein leads to an extended S phase, failure to exit cell cycle, apoptosis, and cell number decrease in mcm5 mutant embryos. However, apoptosis increases only in the retina, tectum, and hindbrain but not in other late-proliferating tissues - around the third day of development - suggesting that different tissues may employ distinct cellular programs in response to MCM5 depletion66. We also identified a downregulation of CDC45 a member of the highly conserved multiprotein complex which interacts with MCM7 and appears involved in mortality/aging, cellular phenotype, and embryo phenotype. The mRNA expression is detected in embryonic tissues and stem cells from neural tube, mesencephalic ventricular zone, telencephalon, diencephalic ventricular zone, metencephalic alar plate and brain, choroid plexus and medulla oblongata and phenotypes associated included cognitive impairment and cognitive decline measurement38. Additionally, we also detected a downregulation of MRE11A gene that encodes a nuclear protein involved in homologous recombination, telomere length maintenance, and DNA double-strand break repair. As part of the MRN complex (a complex of three proteins — Mre11, Rad50 and Nbs1), it interacts with MCM9 and the interaction recruits the complex to DNA repair sites67. A component of the BASC complex (BRCA1-associated genome surveillance complex), at least composed of BRCA1, MSH2, MSH6, MLH1, ATM, BLM, RAD50, MRE11 and NBN68, interacts with herpes simplex virus 1 protein UL1269, and in E4 adenovirus infection is inactivated and degraded by viral oncoproteins, thereby preventing concatenation of viral genomes in infected cells. The MRE11A gene also is associated to increased risk of cancer70,71.
Relevance for clinical surveillance
In this pilot study we highlight the cellular and molecular changes and the potential clinical implications in infants exposed prenatally to the Zika virus, not only in CZS, but also for those without clinical signs - asymptomatic (Figure 8; supplemental Table 7). Indeed, the results found alert for surveillance after a viral exposure, especially in asymptomatic individuals, considering the potential risk of a new Zika virus epidemic3. We identified deregulated genes and pathways involved in developmental disorders and cancer risk in CZS infants (Figure 6; Supplemental Table 8). Several parallelisms exist between development and cancer due to the similarities between normal tissue stem cells and cancer stem/progenitor cells72. Several developmental disorders are associated with congenital malformations and cancer risk and the risk is higher in children with congenital anomalies and some specific genetic syndromes73. Nevertheless, little is known regarding how defects in these genes and signaling pathways can produce congenital developmental abnormalities as well as alterations underlying the development of neoplastic process. Tumorigenesis associated with developmental syndromes is related to several variables such as pathways, genes, and genetic/epigenetic alterations as well as the origin of these mutations (somatic or germinative). However, despite the small number of samples evaluated in this pilot study, the monitoring and screening for pediatric cancer predisposition syndromes74 should not be discarded in CZS infants.
Conclusion
Ocular cells used for the first time as a neuroepithelial model revealed the effect of Zika infection on primordial neuronal cell genes, evidenced by changes in genes associated with embryonic cells. Precisely because this is a pilot study, further studies with a larger sampling should be performed to corroborate these findings. Despite the small number of samples evaluated, the results described here support those previously described in the literature about Zika-host interaction. The changes in the genes validate an association with the gestational period of the occurrence of the infection and testify to the resulting clinical and ophthalmological pathologies. On the other hand, evidence that there is a deregulation in genes related to cell death brings to the fore the concern with the early onset of other associated pathologies. Evidence of deregulated genes associated with cancer raises concerns about the surveillance regarding the appearance of tumors. Here we support the hypothesis that the CNS changes in infants with CZS may be occurring beyond the consequences of the congenital infectious process. Thus, it will be interesting to study whether regulatory changes of the primordial genes of development during embryogenesis, as detected in this pilot study, might contribute to the neuropathological condition observed in this syndrome.
Data Availability
The data for this study have been deposited in the European Nucleotide Archive at EMBL EBI under accession number PRJEB40869. Additional data supporting the findings of this study are available within the article and its Supplemental files or are available from the authors upon request. Due to privacy and consent restrictions, individualized RNAseq data files cannot be uploaded publicly, but might be available to researchers upon request and signature of a Data Use Agreement.
Data availability
The data for this study have been deposited in the European Nucleotide Archive (ENA) at EMBL-EBI under accession number PRJEB40869 (https://www.ebi.ac.uk/ena/browser/view/PRJEB40869). Additional data supporting the findings of this study are available within the article and its Supplemental files or are available from the authors upon request. Due to privacy and consent restrictions, individualized RNAseq data files cannot be uploaded publicly, but might be available to researchers upon request and signature of a Data Use Agreement.
Author contributions
Funding /Support
This pilot study was supported by grants from Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ, Brazil, Proc. n.º 201.779/2017 - PDS/2017), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil -RCNM and GP), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, Brazil, CBB-APQ-03647-16 -RCNM) and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, Brazil, Proc. n.º 2014/06863-3, 2018/18257-1, 2018/15549-1). RH Barbosa was a recipient of a Brazilian FAPERJ senior postdoctoral fellowship.
Tables
Table 1. Descriptive table of expression differentiated of transcripts into ZIKV/CZS and CTRL babies.
Table 2. Descriptive table of expression differentiated of transcripts into asymptomatic ZIKV and CTRL babies.
Supplemental Table 1. Clinical data.
Supplemental Table 2. Deregulated genes in ZIKV/CZS babies, enriched GO interaction network according STRING/KEGG analysis.
Supplemental Table 3. Deregulated genes in Asymptomatic ZIKV babies, enriched GO interaction network according STRING/KEGG analysis.
Supplemental Table 4. Correlation gene expression / Congenital Zika Syndrome phenotype according embryogenesis stages related. Results from Toppgene analysis.
Supplemental Table 5. Correlation gene expression / Congenital Zika Syndrome phenotype according neurological process associated. Results from Toppgene analysis.
Supplemental Table 6. Correlation gene expression / Congenital Zika Syndrome phenotype according ocular phenotype associated. Results from Toppgene analysis.
Supplemental Table 7. Correlation gene expression / Asymptomatic ZIKV phenotype according associated disorders. Results from Toppgene analysis.
Supplemental Table 8. Correlation gene expression / Congenital Zika Syndrome phenotype according associated disorders. Results from Toppgene analysis.
Acknowledgements
We are grateful to Prof. Lemos for helping us with the initial steps in the RNAseq analysis tools and for reviewing of the first versions the manuscript. We thank the Zika Study Group of the Clinical Research Unit from the Universidade Federal Fluminense especially to the patients and their families. We dedicate this work to the Super Special Mom Mrs. Simone Duarte and to our sweet Guilherme (in memoriam).
References



