
ABSTRACT
Background Implementation of Option B+ antiretroviral therapy (ART) has significantly lowered vertical transmission rates but has also increased numbers of HIV-exposed uninfected children (HEU), who remain vulnerable to morbidities. Here, we investigated whether altered immune status in HEU originates in the placenta.
Methods We analyzed T cells from term placentae decidua and villous tissue and paired cord blood from pregnant women living with HIV (PWLWH) who initiated ART late in pregnancy (n=21) with HIV negative controls (n=9).
Results Placentae from PWLWH showed inverted CD4:CD8 ratios and higher proportions of tissue resident CD8+ T cells in villous tissue relative to control placentae. CD8+ T cells in the fetal capillaries, which were of fetal origin, positively correlated with maternal plasma viraemia prior to ART initiation, implying that imbalanced T cells persisted throughout pregnancy. Additionally, the expanded memory differentiation of CD8+ T cells was confined to the fetal placental compartment and cord blood, but was not observed in the maternal decidua.
Conclusion T cell homeostatic imbalance in the blood circulation of PWLWH is reflected in the placenta. The placenta may be a causal link between HIV-induced maternal immune changes during gestation and altered immunity in newborn infants in the absence of vertical transmission.
Lay Summary The effective prevention of HIV transmission during pregnancy with the rollout of antiretroviral therapy (ART) has resulted in increased numbers of HIV-exposed uninfected children (HEU). These children are vulnerable to infections and health problems and they have altered cellular immune systems at birth. We investigated whether these immune alterations may originate in the placenta, as this fetal organ maintains life during pregnancy. Do immune alterations in the newborn child originate in the placenta? After collecting placentae at term from pregnant women living with HIV (PWLWH), who started ART in the third trimester (n=21) and from HIV negative controls (n=9), we isolated T cells from dissected placental tissue and matching cord blood. Placentae from PWLWH showed inverted CD4:CD8 ratios in the placenta and cord blood with higher numbers of CD8+ T cells in the fetal part of the placenta. These CD8+ T cells mirrored events in the blood circulation of the mother and the altered balance of T cell immunity in the PWLWH was reflected in the placenta. Accordingly, the placenta may be a pivotal link between HIV-induced maternal immune changes and altered immunity in newborn infants in the absence of vertical transmission.
Background
In adults, HIV causes severe immune dysregulation, characterized by systemic depletion of CD4+ T cells, increased HIV-1 specific CD8+ T cells, inflammation and a progressive failure of the immune system[1–3]. Initiation of ART has been shown to augment HIV-specific CD4+ T cell responses, but normalization of the CD4:CD8 T cell ratio does not occur in a large proportion of HIV positive individuals[4]. In pregnant women living with HIV (PWLWH), pregnancy is not associated with HIV disease progression, although it has been associated with functional impairment of systemic effector T cells and an overall altered response against HIV control[5,6].
Placentae from PWLWH exhibit increased signs of inflammation and injury affecting maternal vasculature and circulation[7,8]. Although the maternal and fetal circulation within the placenta takes place in distinct compartments, there is evidence that maternal HIV infection impacts the fetal immune system. HIV-unexposed uninfected children (HUU) have an almost completely naïve T cell repertoire, but at birth HIV-exposed uninfected children (HEU) can have increased proportions of differentiated immune cells suggestive of antigen experience in utero[9,10]. This scenario raises the question of how HIV exposure in utero potentially creates perturbations of immunity in HEU[11,12].
In healthy human pregnancies, T cells in the placenta have been shown to constitute approximately 5-20% of total leukocytes in the decidua and this proportion increases up to over 40% at term[13,14]. CD8+ T cells are the most abundant subset and are important in promoting embryo implantation and providing immune protection against infection[15,16]. CD4+ T cell subsets have a number of key roles including the establishment and maintenance of fetal-maternal (FM) tolerance[17,18]. Fetus-specific CD4+ Tregs have been shown to migrate towards the FM interface where they are enriched to guard against breakdown in FM tolerance[18]. In contrast, the presence of CD8+ T cells in the villous tissue is often associated with villitis of unknown etiology (VUE), an inflammatory state of the placenta characterized by the infiltration of maternal CD8+ T cells and macrophages into the villi[19,20].
To test the hypothesis that maternal HIV infection is associated with disruption of T cell homeostasis in the placenta and cord blood from HEU newborns, we examined term placentae from PWLWH from a randomized trial in pregnant mothers initiating dolutegravir versus efavirenz-containing therapy in the third trimester (DolPHIN-2: NCT03249181)[21], as well as pregnant women not living with HIV (PWNLHIV), serving as controls. We show that placentae from PWLWH have inverted CD4:CD8 ratios with higher CD8+ T cells in villous tissue relative to control placentae contributing to T cell homeostatic imbalance in the placenta at birth.
Methods
Cohort
We included 21 placentae with 9 paired cord blood samples from PWLWH and HEU and 9 placentae from PWNLHIV with 5 cord blood samples from HUU in this study. The PWLWH group was nested in the DolPHIN-2 study and recruited from the Gugulethu Community Health Centre, Cape Town[21]. HIV negative controls were enrolled nearby from the Khayelitsha Site B Midwife Obstetric Unit, Cape Town. All placentae were from term deliveries (>37 weeks’ gestation).
Clinical Data collection
As part of DolPHIN-2, maternal systemic CD4 T cell counts were measured at ART initiation and plasma viral load (VL) copies were measured at screening, enrolment and ART initiation (visit 1), one week after ART initiation (visit 2), four weeks after ART initiation (visit 3), thirty-six weeks’ GA (visit 4) and approximately ± 14 days after delivery (visit 5). The level of detection was 50 copies per ml[21].
Placenta and cord blood processing
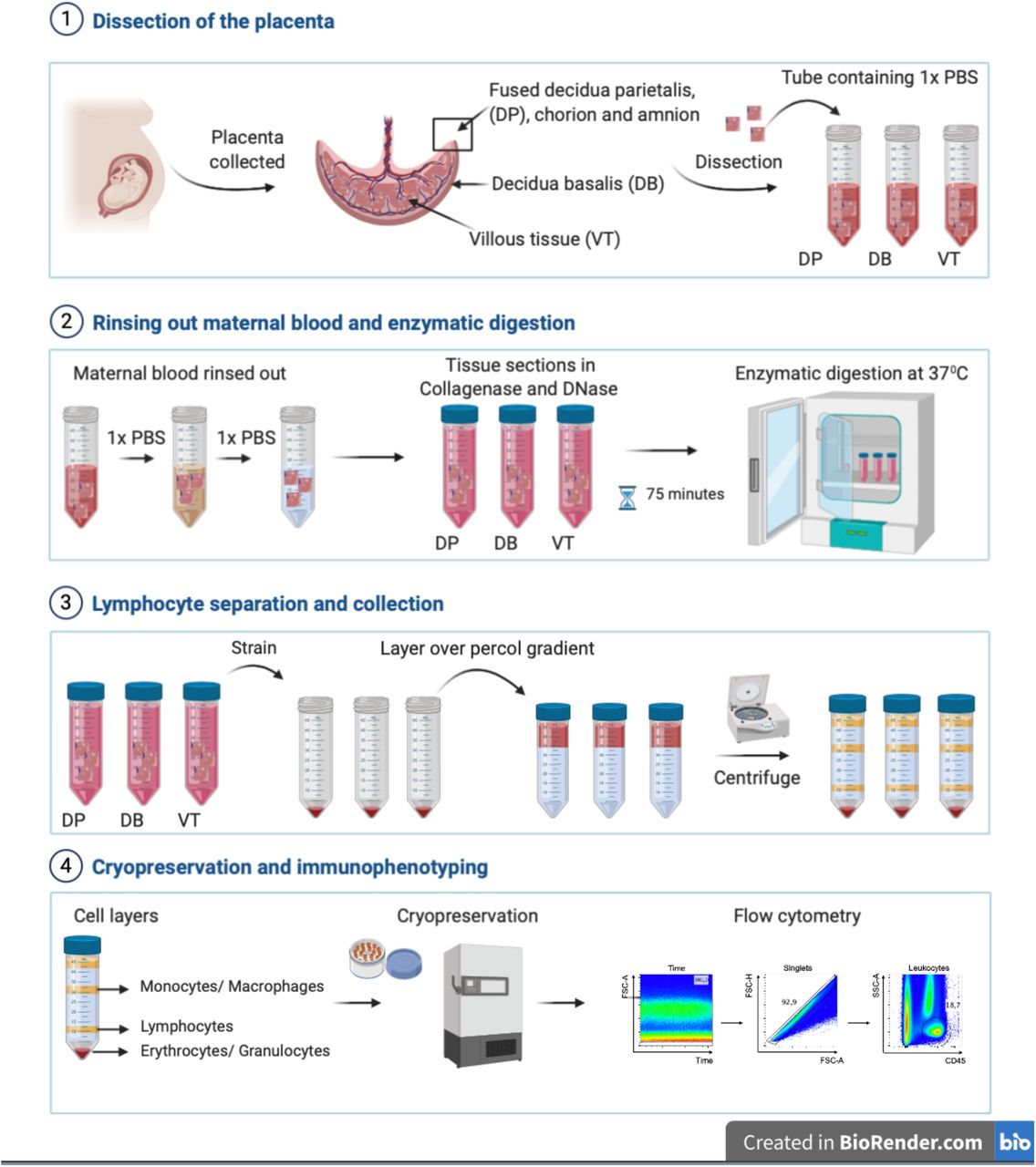
Cells were isolated from each placenta as previously described[22] and illustrated in Figure 1. Placentae were collected in RPMI 1640 supplemented with 10% fetal calf serum (FCS) and penicillin/streptomycin at room temperature and processing was performed within six hours of delivery. Briefly, each placenta was macroscopically inspected and dissected to obtain the decidua parietalis, basalis and villous tissue. The sections were rinsed in phosphate-buffered saline (PBS) to remove maternal blood. Enzymatic lymphocyte isolation was performed using 0.2% Collagenase I and 0.02% DNAse I in RPMI 1640 at 37°C for 75 minutes. We then obtained the lymphocyte fraction following Percoll density centrifugation at the 70-45% interface and incubated the cells with violet amine reactive viability dye (VIVID, Thermofisher) according to manufacturer’s instructions. The cells were then fixed using BD FACS™ lysing solution, cryopreserved in 90% FCS - 10% Dimethyl sulfoxide (DMSO) and stored in liquid nitrogen until analysis. Cord blood mononuclear cells were isolated on Ficoll, fixed and cryopreserved in the same fashion as placental lymphocytes.

Stepwise isolation of lymphocytes from the human placenta decidua parietalis (DP), decidua basalis (DB) and villous tissue (VT): (1) dissection of the whole placenta; (2) multiple rounds of maternal blood rinsing to avoid contamination followed by enzymatic digestion of each dissected tissue; (3) lymphocyte separation obtained by Percoll density centrifugation; (4) cryopreservation of fixed cells and immunophenotyping using flow cytometry.
Placenta pathology
Placentae were fixed in 10% buffered formalin prior to histopathology examination at Red Cross Memorial Children’s Hospital, Cape Town, South Africa. Specimens were macroscopically examined and samples from the umbilical cord, placental membranes and placental disk were obtained based on the Amsterdam Placental Workshop Group Consensus Statement.[23] Briefly, four blocks were prepared from each placenta; including a roll of the placental membranes, two cross sections of the umbilical cord; and full-thickness sections of the placental parenchyma including one from the cord insertion. Tissue sections were 3-5μm thick and were stained with haemotoxylin and eosin for routine histological examination. Placentae were examined for features of prolonged meconium exposure, chorioamnionitis with or without a maternal or fetal inflammatory response[24,25], maternal vascular malperfusion (MVM) with decidual vasculopathy, and in the absence of decidual vasculopathy, MVM was indicated by a small placenta for gestational age accompanied by at least two villous changes (villous infarct, retroplacental haematoma, accelerated villous maturation or distal villous hypoplasia). In the case of normal placenta weights for gestational age, at least 3-4 villous features were noted to be placed in the category of MVM[26]. Chronic deciduitis was defined as abnormal infiltration of lymphocytes and plasma cells in the decidua[23].
Flow cytometry
Placental and cord blood cells were labeled with fluorochrome-conjugated monoclonal antibodies: CD3 (Clone UCHT1), CD4 (Clone SK3), CD8 (Clone SK1), CD45RA (Clone H100), CD28 (Clone CD28.2), CD14 (Clone MHCD1417) and CD45 (Clone MHCD4530). Samples were acquired using an LSR II flow cytometer (BD Biosciences). Total CD4+ and CD8+ T cells were expressed as a proportion of CD3+ T cells gated from the viable CD45+ CD14-cells (Supplementary Figure 1).
Immunohistochemistry
Formalin fixed paraffin embedded (FFPE) placenta tissue blocks were cut into 5 μM sections and stained with CD8 (Clone C8/144B), with tonsillar tissue serving as a control. Briefly, the slides were baked overnight at 56°C and rehydrated in xylene followed by varying concentrations of alcohol and then incubated in 3% hydrogen peroxide. Heat-mediated antigen retrieval was performed using an EDTA buffer (pH9). The slides were then incubated with 1% Bovine Serum Albumin (BSA) and stained with the primary antibody anti-CD8. The images were acquired on Zeiss Axioskop 200 upright Fluorescence microscope with an AxioCam high resolution colour (HRC) camera.
Fluorescence in situ hybridization (FISH)
We used five placental samples from male fetuses to identify the origin of the infiltrating lymphocytes by FISH. Briefly, the slides were baked at 90°C for 15 minutes, deparaffinized in xylene, dehydrated in 100% ethanol and then placed in 10mM Citric Acid (pH 6.0). The slides were then dehydrated in varying concentrations of ethanol (70%, 85% and 100%). We then applied a working solution of DXZ1/DYZ3 (Abbott Laboratories, Des Plaines, IL, USA) to the target areas, co-denatured with a ThermoBrite (Abbott Laboratories) and hybridized overnight at 37°C. The slides were then counter stained with 4’-6’-diamidino-2-phenylindole (DAPI) (Vector Laboratories). Tissue samples were scanned and the qualitative result was determined based on observed signal patterns by CytoVision (Leica Biosystems, Germany).
Statistics
All flow cytometry data were analyzed using FlowJo version 10 (Treestar). Statistical analyses were performed using Prism version 8 (Graphpad Software, San Diego, CA), STATA version 12.0 (Stata Corporation, College Station, Texas, USA) or R[27]. Immunohistochemistry tissue cell counts for each tissue section were obtained by counting the total number of positive CD8+ T cells on the immunostained slides (40x magnification) using Image J Fiji version 2 (WS Rasband, National Institute of Health, Bethesda, MD). Tests of significance were performed using Mann-Whitney U and Kruskal-Wallis tests for intergroup comparisons. The associations between cell proportions and viral load or CD4 T cell counts were assessed using simple linear regression. All bivariate analyses including maternal and infant characteristics, and placental pathology stratified by HIV-exposure or by ART regimen were compared using Chi2 or Fisher’s exact test and Wilcoxon rank-sum tests.
Study approval
The study protocol, informed consent forms and all data collection tools were approved by the University of Cape Town, Faculty of Health Sciences Research Ethics Committee. Written and signed informed consent was obtained from all participants, including collection of placentae prior to study inclusion.
Results
Participant Characteristics
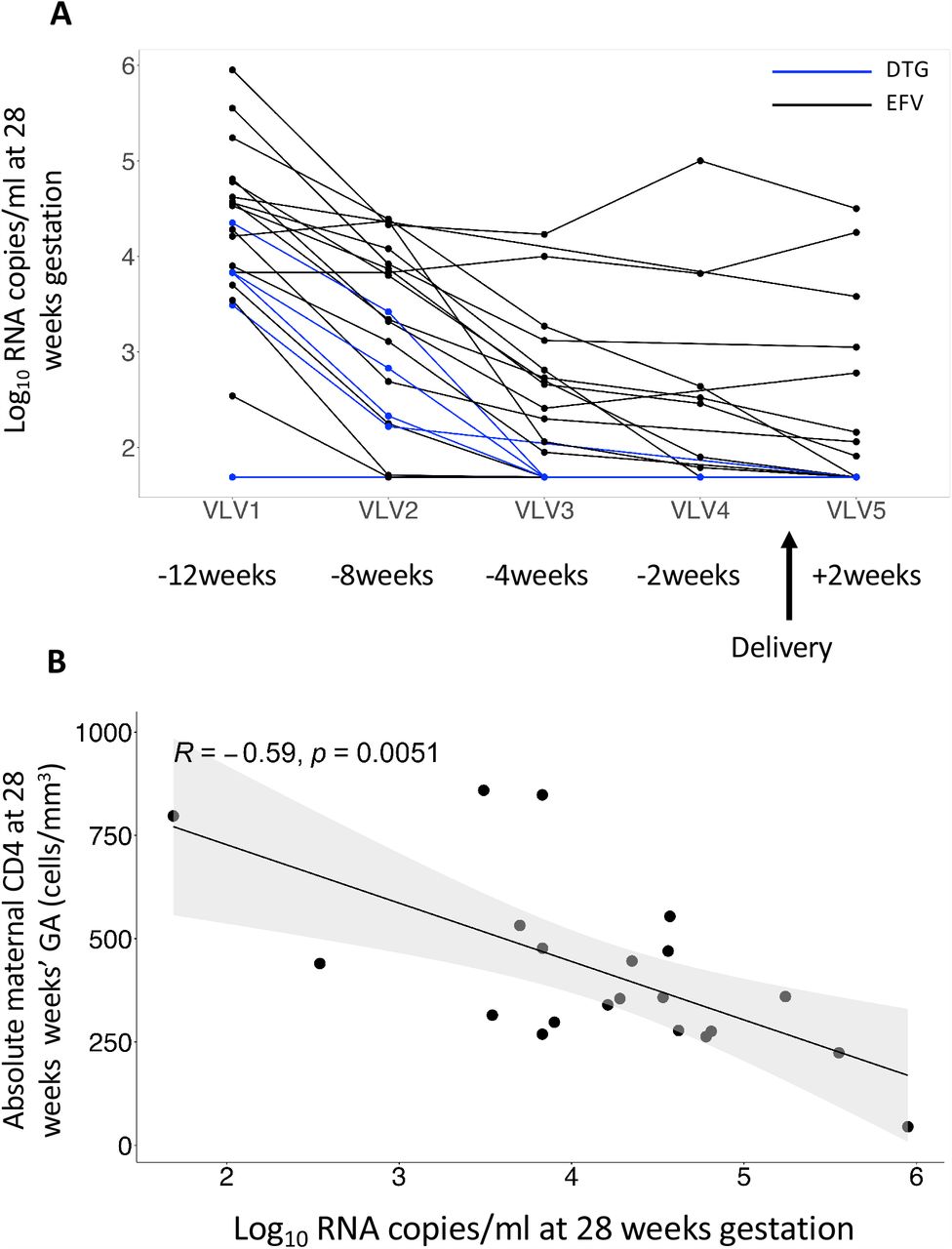
Maternal and newborn infant characteristics are shown in Table 1. No differences in maternal age were noted between PWNLHIV and PWLWH at enrolment, with the median gestational age (GA) being 30 weeks and 28 weeks between the groups. PWLWH were more likely to be multigravida (p=0.003). Median GA at delivery was 40 weeks in PWNLHIV and 39 weeks in the PWLWH (p=0.03) and there was a tendency toward lower birthweight in HEU (p=0.07). Among the 21 PWLWH, 16 (76.2%) were randomised to receive efavirenz (EFV + TDF + 3TC) and 5 (23.8%), dolutegravir (DTG + TDF + 3TC), shown in Supplementary Table 1. Median CD4 count at ART initiation was 358 cells/mm3 (IQR 278 – 477), with no difference between ART groups (Supplementary Table 2). The median VL at ART initiation was 4.54 log10 RNA copies/ml (IQR 3.85 – 4.80) in the EFV group versus 3.83 log10 RNA copies/ml (3.49 – 3.83) in the DTG group, with a combined viraemia of 4.28 log10 RNA copies/ml (Supplementary Table 2). Both ART groups were on treatment for a median of 84 days IQR (44 - 105) and women in the DTG arm achieved viral suppression at a faster rate (cut off ≤ 50 copies/ml or 1.69 log10 RNA copies/ml) at 4 weeks versus 2 weeks after delivery (Supplementary Table 2 and [21]). For the purposes of this study, we combined the placentae from two ART groups, as there were only 5 collected from the DTG arm.
Maternal and infant characteristics
Placental characteristics and pathology at delivery
Placental weight is altered by HIV
Table 2 shows placenta characteristics and pathology stratified by HIV status. PWNLHIV had significantly larger placentae (468g, IQR 426 - 533) compared to the PWLWH (394g, IQR 343 - 469; p=0.02), with 38% of placentae in PWLWH being <10th percentile weight-for-gestation compared to 0% in PWNLHIV. These differences were also reflected the in the fetal-placental (FP) ratios, where all cases with FP ratios > 90th percentile were in the PWLWH cases (p=0.02). Placental histopathology identified 2 cases (9.5%) of chronic deciduitis and 6 cases (28.6%) of MVM, all in the PWLWH. There were no significant differences in the incidence of meconium exposure and chorioamnionitis between PWLWH and PWNLHIV, and there was no evidence of villitis of unknown etiology (VUE) in any of the placentae. There were no significant differences in the placental weight, FP ratio and placental pathology between the two ART groups (Supplementary Table 3).
HIV infection during pregnancy alters placental CD4+ and CD8+ T cell proportions
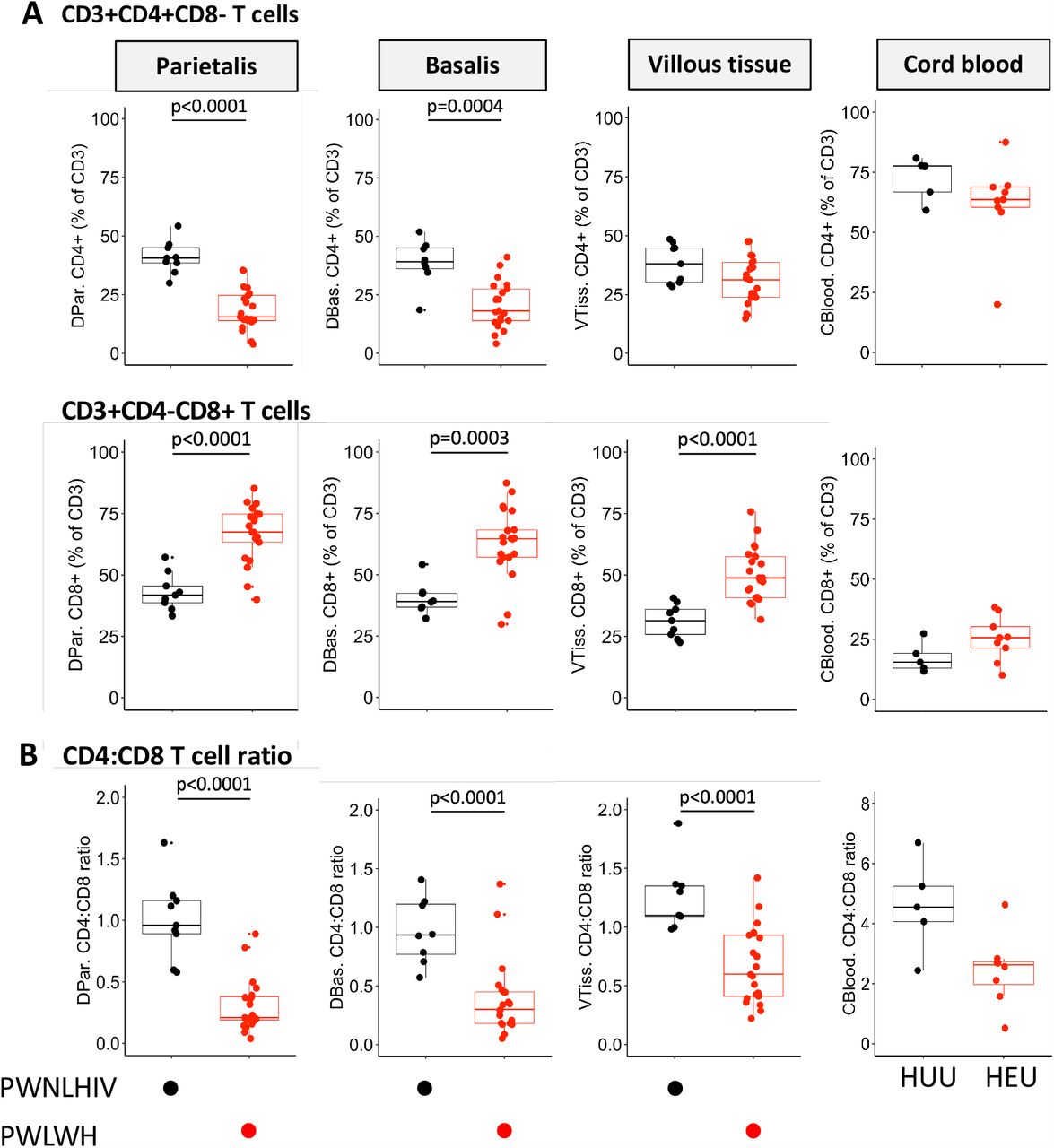
Figure 2A shows significantly lower proportions of CD4+ T cells in decidua parietalis and basalis, but not in the villous tissue, comparing placentae from PWLWH with PWNLHIV. The proportion of CD8+ T cells was significantly increased (Figure 2A) in all three placental compartments resulting in significantly lower CD4:CD8 T cell ratios in the three placental tissues from PWLWH (Figure 2B). Notably, the inverted CD4:CD8 ratio in villous tissue was due to the increased CD8+ T cells in the villous tissue. The inverted CD4:CD8 ration was partially reflected in cord blood from HEU (Figures 2B). No differences were identified in T cell proportions when stratified by the different ART regimens (Supplementary Figure 2).


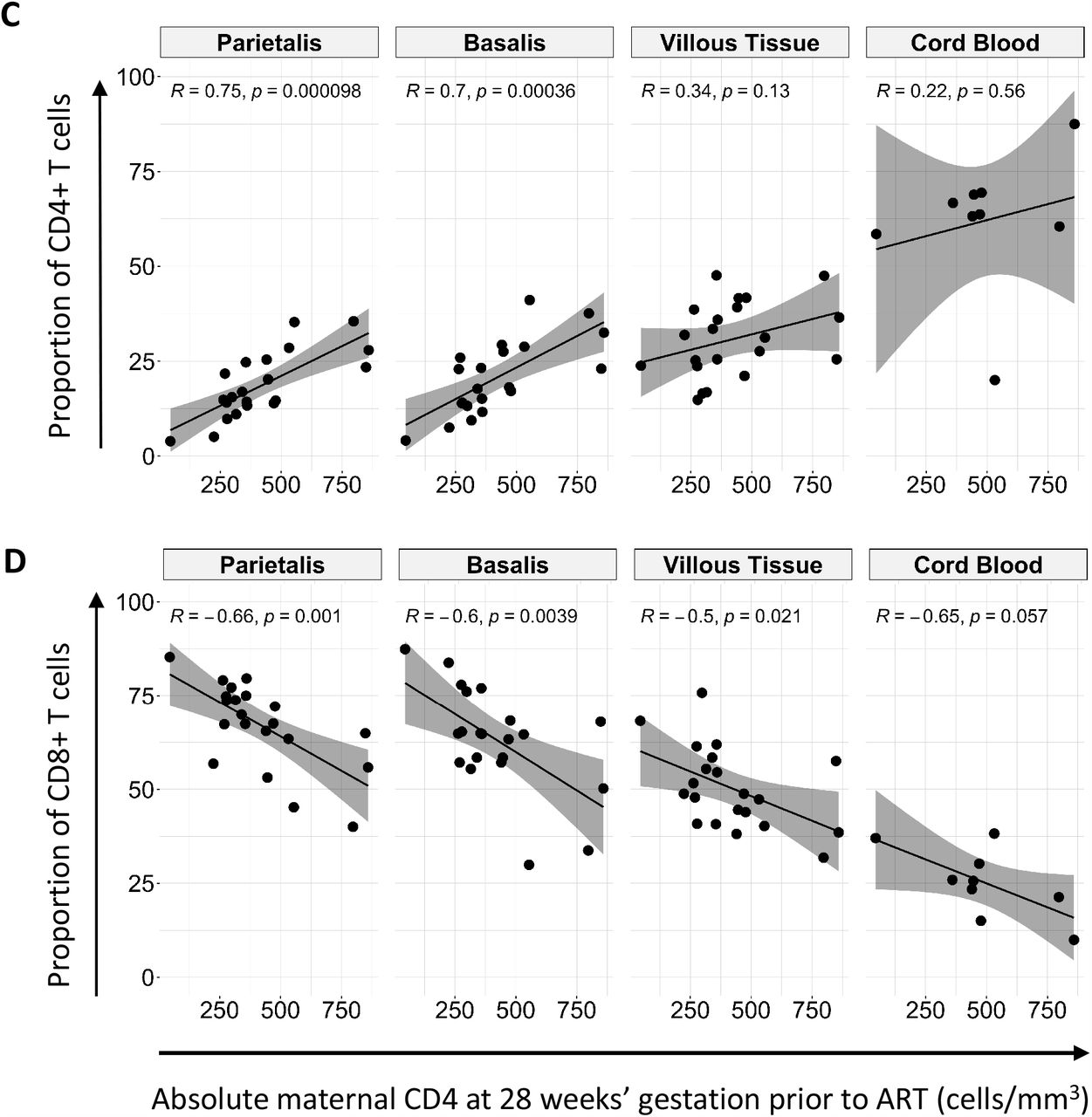
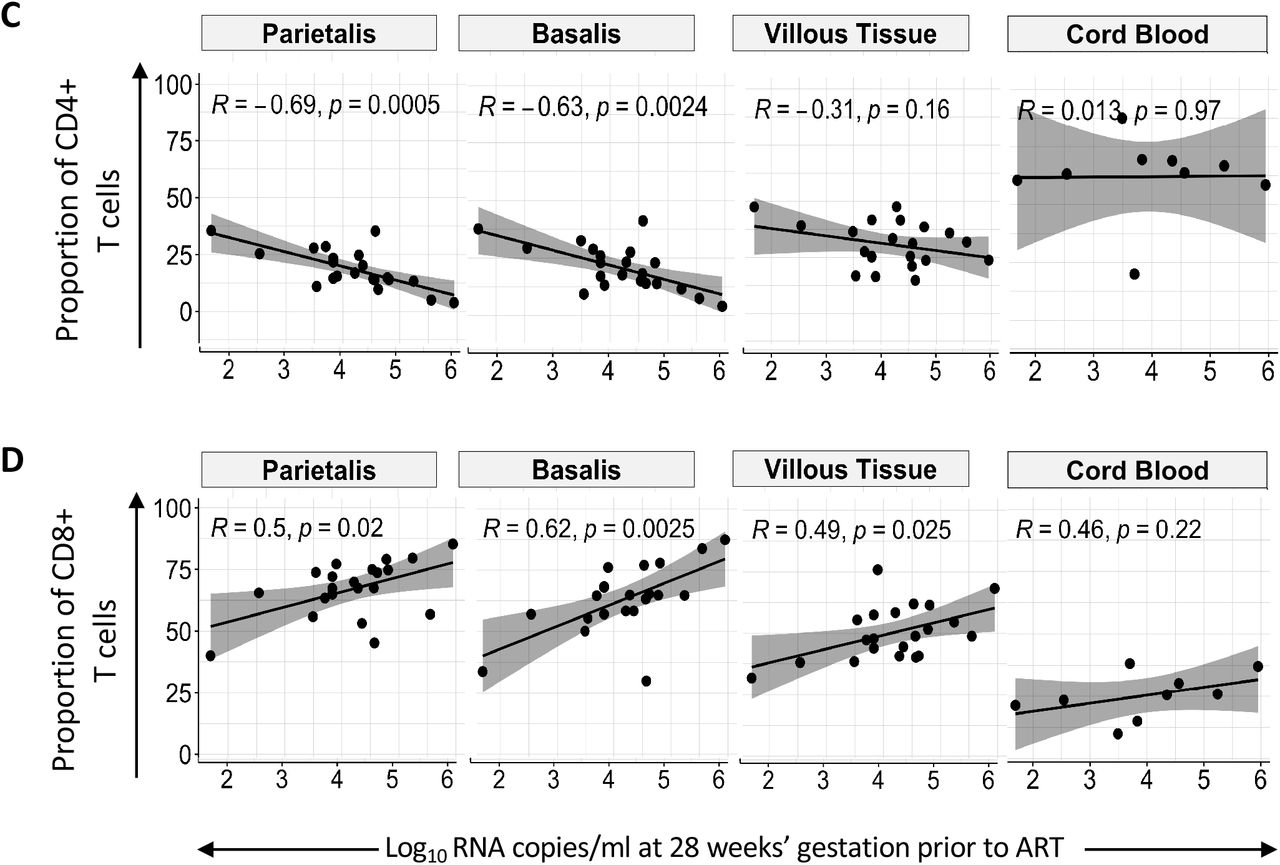
(A) Box plots (showing medians and interquartile ranges) of CD3+CD4+CD8-T cells and CD3+CD4-CD8+ T cell proportions isolated from the decidua parietalis, decidua basalis, villous tissue and cord blood from Pregnant Women not living with HIV (PWNLHIV) and Pregnant Women living with HIV (PWLWH) and HIV unexposed uninfected (HUU) and HIV exposed uninfected (HEU) cord bloods. (B) Box plots (showing medians and interquartile ranges) of CD4:CD8 T cell ratios in the decidua parietalis, decidua basalis, villous tissue and cord blood from Pregnant Women not living with HIV (PWNLHIV) and Pregnant Women living with HIV (PWLWH) and HIV unexposed and uninfected (HUU) and HIV exposed uninfected (HEU) cord bloods. Tests of significance were performed using the Mann-Whitney U test. (C) Correlation plots between the absolute maternal CD4 count at 28 weeks’ gestation prior to ART initiation and the proportion of CD4+ T cells isolated from the decidua parietalis, basalis, villous tissue and cord blood. Statistical analysis was performed using the Spearman rank test and the grey shaded areas represent the 95% confidence intervals. (D) Correlation plots between the absolute maternal CD4 count at 28 weeks’ gestation prior to ART initiation and the proportion of CD8+ T cells isolated from the decidua parietalis, basalis, villous tissue and cord blood. Statistical analysis was performed using the Spearman rank test and the grey shaded areas represent the 95% confidence intervals.
Maternal absolute peripheral blood CD4 T cell counts, measured pre-ART at a median of 28 weeks’ gestation, positively correlated with the proportion of CD4+ T cells in the decidua and negatively correlated with the proportion of CD8+ T cells in decidua and villous tissue (Figure 2C and D). A similar trend, without significance, was observed in cord blood. Thus, the inverted placental tissue CD4:CD8+ T cell ratios appeared to reflect the maternal peripheral immune status but was mirrored to a lesser extent in the cord blood of HEU. This correlation was temporally dissociated, where correlations were made between blood measured at 28 weeks and placentae measured at 38-40 weeks of gestation, suggesting that inverted T cell ratios persisted throughout pregnancy.
Maternal viral load prior to ART initiation correlates with placental, but not cord blood, CD4+ and CD8+ T cell proportions
Maternal plasma viraemia dropped precipitously over time from enrolment to delivery (−12 weeks beforehand), where PWLWH receiving DTG decreased at a faster rate (Figure 3A)[17]. As expected, the enrolment plasma viraemia, ranging from 1.69 - 6.0 log10 RNA copies/ml, significantly inversely correlated with the absolute maternal peripheral blood CD4+ T cell count determined pre-ART, at enrolment (Figure 3B). Interestingly, pre-ART viremia also showed a significant negative correlation with proportions of CD4+ T cells in the decidua parietalis and basalis (Figure 3C) and a positive correlation with CD8+ T cells in the decidua parietalis, basalis and villous tissue at delivery (Figure 3D). The association between maternal viral loads over time and the proportions of decidual CD4+ and CD8+ T cells were maintained at −8 weeks and −4 weeks before delivery for CD4+ T cells (Supplementary Figure 3) and up to −8 weeks before delivery for CD8+ T cells (Supplementary Figure 4). Maternal viremia did not correlate with the proportion of T cells in the cord blood (Figure 3C and 3D), suggesting that maternal VL (pre- and post-ART initiation) can influence the homeostatic balance of T cells in the placenta, but not in the “newborn” immune compartment.


(A) Line plot depicting participant viral load trajectories over time at enrolment and ART initiation (VLV1, 12 weeks before delivery), one week after ART initiation (VLV2, 8 weeks before delivery), four weeks after ART initiation (VLV3, 4 weeks before delivery), thirty-six weeks’ GA (VLV4, 2 weeks before delivery) and +/- 14 days after delivery (VLV5, +/- 2 weeks after delivery). The women were randomised to receive efavirenz (EFV + TDF + 3TC) (denoted in black) and dolutegravir (DTG + TDF + 3TC) (denoted in blue). (B) Correlation plot between maternal systemic absolute CD4 counts at enrolment prior to ART initiation with maternal viral load at enrolment and ART initiation (28 weeks GA). Statistical analysis was performed using the Spearman rank test and the grey shaded area represents the 95% confidence intervals. (C) Correlation plots between CD4+ T cell proportions in the placenta and maternal viral load at enrolment and ART initiation (28 weeks GA) in the decidua parietalis, basalis, villous tissue and cord blood. Statistical analysis was performed using the Spearman rank test and the grey shaded area represents the 95% confidence intervals. (D) Correlation plots between CD8+ T cell proportions in the placenta and maternal viral load at enrolment and ART initiation (28 weeks GA) in the decidua parietalis, basalis, villous tissue and cord blood. Statistical analysis was performed using the Spearman rank test and the grey shaded area represents the 95% confidence intervals.

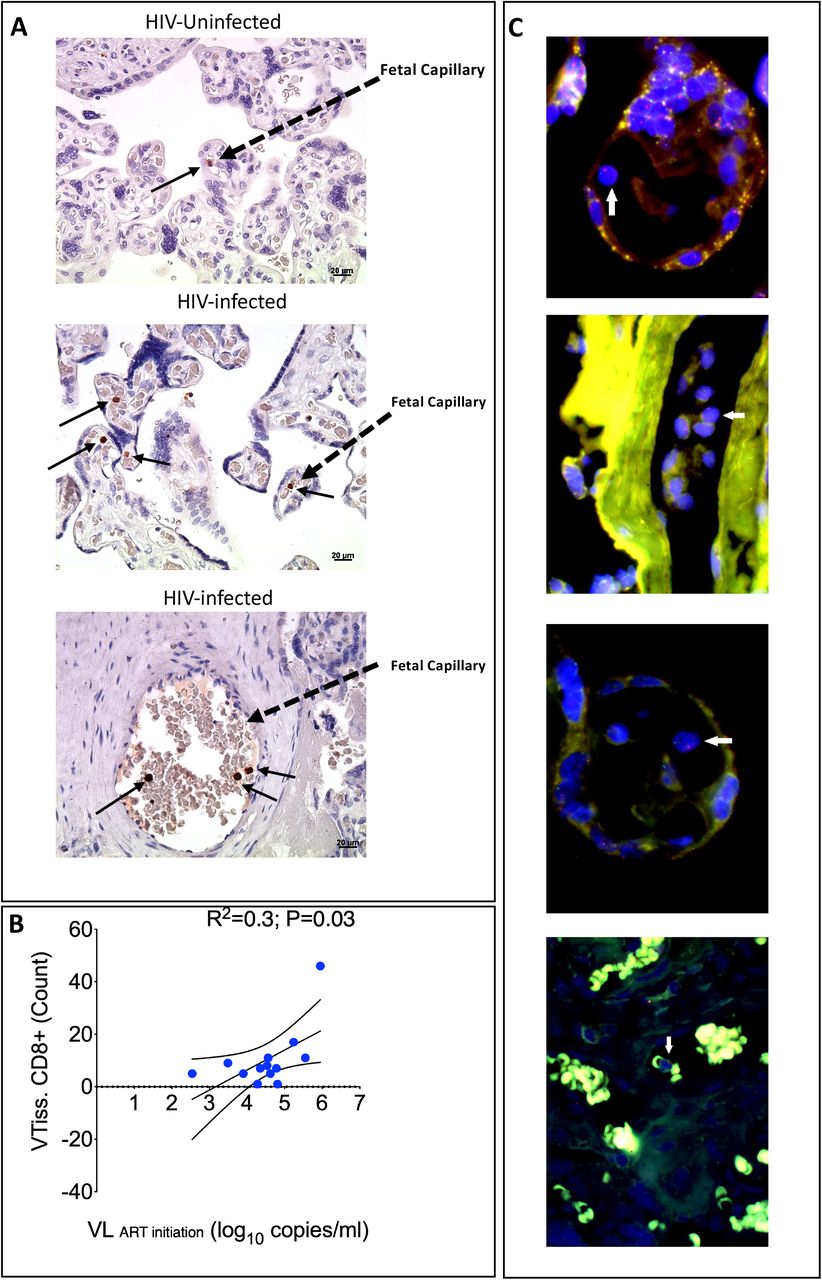
(A) Representative immunohistochemical stained images of CD8+ T cells in villous tissue sections denoted in brown dots and black arrows in the villi of placentae from HIV-infected and –uninfected mothers (40x magnification). (B) Correlation plot between the density of tissue-bound CD8+ T cells in the villi and maternal viral load at ART initiation (pre-ART). Statistical analysis was performed using the Spearman rank test and the black curved lines represent the 95% confidence intervals. (C) Representative Fluorescence in situ Hybridization (FISH) images of lymphocytes (white arrows) in the villous tissue from placentas from male infants. The X chromosome is denoted in green and Y chromosome denoted in red (digitally scanned slides).
Immunohistochemistry confirms anatomical location and fetal origin of CD8+ T cells in villous tissue
Analysis of placental villi from 13 PWLWH and 3 PWNLHIV controls showed the presence of CD8+ cells in the fetal capillaries (Figure 4A). The numbers of CD8+ T cells positively correlated with pre-ART maternal viremia (Figure 4B), confirming the flow cytometric analysis (Figures 2A, 3D). Using FISH to detect X and Y chromosomes in cells located within the villi from 5 placentae with male births, we confirmed the presence of male (fetal) cells in fetal capillaries (Figure 4C). Thus the immunohistochemistry analysis confirms the anatomical location and fetal origin of the increased proportions of CD8+ T cells in villous tissue. The fetal (male) origin of the CD8+ T cells in placental villi is consistent with the absence of VUE, a lesion that is characterized by maternal immune infiltrates[20,28].
HIV exposure increases differentiation of CD8+ T cells in placental villi and fetal cord blood, but not in the maternal placental compartments
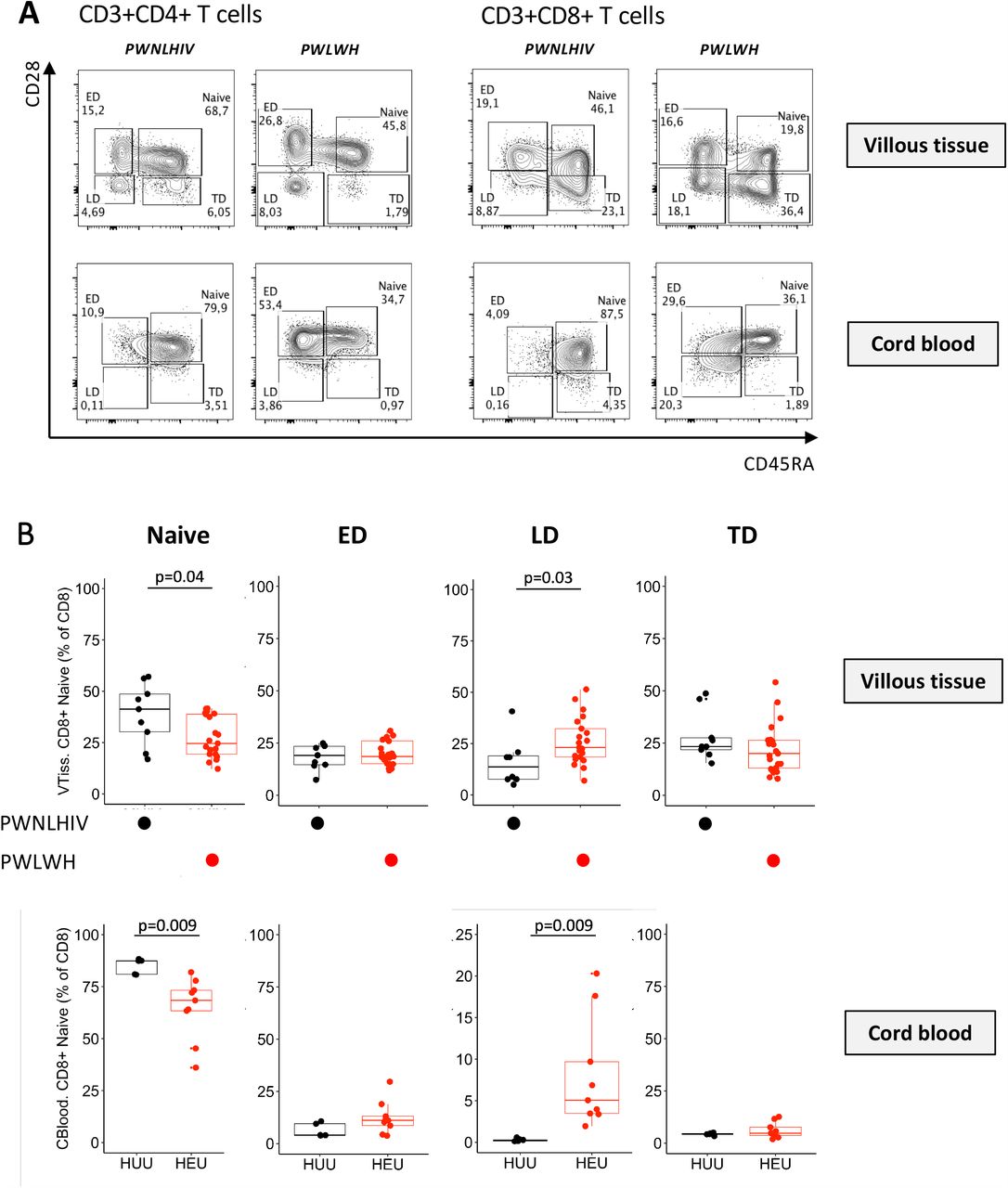
CD45RA and CD28 was used to identify the proportions of naïve (CD45RA+CD28+), early differentiated (ED, CD45RA-CD28+), late differentiated (LD, CD45RA-CD28-) and terminally differentiated (TD, CD45RA+CD28-) memory CD4+ and CD8+ T cells from villous tissue and matching cord blood (Figure 5A). We observed significantly lower proportions of naïve CD8+ T cells and significantly higher proportions of LD CD8+ T cells in villous tissue and cord blood of HEU compared to HUU (Figure 5B). The CD4+ T cells in the villous tissue were predominantly of a naïve and ED phenotype while the cord blood cells were predominantly naïve. There were no significant differences in CD4+ T cell differentiation state based on HIV-exposure (Supplementary Figure 5). In addition, no significant differences in the stage of CD4+ and CD8+ T cell differentiation in decidua parietalis and decidua basalis was observed between the HIV groups (Supplementary Figure 6). Thus, the increased differentiation state of CD8+ T cells is confined to the fetal placental and fetal cord blood compartments and not observed in the maternal placental compartments. Interestingly, there was no correlation between maternal pre-ART VL and memory stage of CD8+ T cells in placental villous tissue and cord blood (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Representative flow cytometry contour plots of Naïve, Early differentiated (ED), Late differentiated (LD) and Terminally differentiated (TD) CD4+ T cells and CD8+ T cells in the villous tissue (VT; upper panel) of placentae from PWNLHIV and PWLWH and cord blood from HIV unexposed and uninfected (HUU) and HIV exposed uninfected (HEU). (B) Box plots (showing medians and interquartile ranges) of CD8+ Naïve, Early differentiated (ED), Late differentiated (LD) and Terminally differentiated (TD) T cells in the villous tissue and cord blood from HIV unexposed and uninfected (HUU) and HIV exposed uninfected (HEU). Tests of significance were performed using the Mann-Whitney U test.
Discussion
We present data from a unique cohort of PWLWH who initiated ART late in pregnancy and show that maternal HIV infection has a clear impact on T cell subsets in the decidua, villous tissue and cord blood. As the decidua and decidual immune cells are of maternal origin, it may not be surprising to find such a footprint[29]. Maternal HIV infection likely affects and kills maternal decidual CD4+ T cells and fewer peripheral blood T cells may traffic to decidual tissue; chemokine gradients have been shown to play a key role in the trafficking of maternal T cells into the decidua during pregnancy[30]. We show that in contrast to the decidua, the inverted CD4:CD8 ratio in the fetal villous tissue was largely due to an increased proportion of CD8+ T cells and not, as observed in the other tissues, due to a decrease in CD4+ T cells. These CD8+ T cells were of an early-late differentiated phenotype, suggestive of previous antigen experience[31,32].
The human placenta has two circulatory compartments: the utero-placental unit for the trafficking of maternal blood and the feto-placental unit for the fetal blood circulation[33,34]. Approximately 40% of the total fetal-placental blood volume exists as a reservoir in the placenta[34]. The cells we characterised in the villous tissue and cord blood are from the feto-placental unit and these villous tissue resident cells are likely predominantly from the placental reservoir. A key question is whether the increased CD8+ T cell differentiation in placental villi and cord blood is due to direct exposure to HIV antigens, presence of other pathogens in placental or fetal compartments (e.g. CMV) or increased levels of other non-infectious inflammatory cues in placentae of PWLWH. There is evidence that despite separation of fetal and maternal circulations, viral particles, structural and core HIV proteins have been shown to cross the placental barrier in the absence of fetal infection leading to altered immune profiles in HEU infants[35,36]. A number of studies have also described HIV-specific T cell responses in HEU infants, postulated to have been primed by exposure to HIV antigens in utero[11,37,38]. The magnitude of these responses is greatest soon after birth but are not detected in older HEU children, suggesting that in the absence of continuous exposure these responses wane[10,38]. The lower proportion of naïve cells and increased memory T cells reflected in villous tissue and cord blood mirror previous studies; HEU infants have been shown to have reduced CD4+ T cell numbers and increased CD8+ T cells compared to HUU infants at birth[39,40]. HEU infants have also been shown to have lower naïve cells thought to be due to thymic involution and frequent stimulation and expansion of the antigen-specific T cells in an effort to regenerate the T cell pool[41–43]. Whether the same findings would be recapitulated in PWLHIV with preconception suppressed viral loads is unknown.
Interestingly the elevated proportions of CD8+ T cells found embedded in the placental villous tissue from PWLHIV were shown to be sequestered within the fetal capillaries. Furthermore, these CD8+ T cells within villous tissue were proportional to maternal viremia. The absence of overt VUE corroborates the finding that expanded CD8+ T cell fractions are of fetal and not maternal origin. Previous studies have suggested that the presence of T cells in the villi during normal pregnancy reflect VUE[20]. However, there is emerging evidence, using IHC staining of villi sections from early elective termination placentae, of the presence of CD45+ αβ T cells, although these cells were undetectable at term[44]. In a separate study, using single cell transcriptomic analysis in placentae from women with or without preterm labour, where there were no reported maternal infections, and villous tissue was shown to contain a mixture of fetal and maternal immune cells[45]. We cannot discount the possibility that there was also a mix of maternal and fetal CD8+ T cells in placental villi in our study.
Of particular note, women were ART naïve during the first and second trimester and it is likely that prolonged HIV exposure may have contributed to altered placental development and the significantly lower placental weight observed. Interestingly, all cases of maternal vascular malperfusion (MVM) were reported in placentae from HIV-infected women and possibly reflects placental injury affecting maternal vasculature and perfusion and increasing the risk of an adverse birth outcome[26]. We have previously reported on MVM in placentae from PLWHIV on long-term ART, an incidence of about 27% overall, similar to Kalk et al. [7,46]. It is likely that HIV and/or ART exposure alters factors involved in vascular development, resulting in placental insufficiency and increased risk of adverse birth outcomes[47].
In conclusion, we provide evidence that in utero exposure to HIV results in an altered immune footprint in both the utero-placental and feto-placental compartments. Despite the initiation of ART in the third trimester, resulting in either full or partial maternal viral suppression by the time of delivery, there was a significant imbalance in term placental T cell homeostasis and to a lesser degree in the cord blood. Our data suggests that the placenta may be a causal link between maternal and neonatal T cell perturbations previously observed in HEU newborns.
Data Availability
All data that was used in this manuscript is available from Clive Gray (clive.gray{at}uct.ac.za)
Author contributions
NMI, ML, SK, LM, HBJ, CMG: Conceptualization and design of the study
NMI, TT: Panel design, sample preparation and development of methods
KP: Histopathology scoring and interpretation
NMI, TRM and SD: Statistical analysis
NMI, TT, KP, TM, LM, EAE, RC, HBJ, CMG: Writing the manuscript
Funding
This research was supported by a Fellowship to NMI from the AXA Research Fund, Paris. The content is solely the responsibility of the authors and does not necessarily represent the official views of those of the AXA Research Fund. The research is part of the DOLPHIN-2 clinical trial sponsored by UNITAID (ClinicalTrials.gov NCT03249181).
Supplementary Figures
Supplementary Figure 1: Gating strategy to delineate CD3+ T cells
Representative flow cytometry dot plots depicting the gating strategy to delineate CD4+ and CD8+ T cells gated from a live CD45+ CD14-CD3+ population in the placenta.
Supplementary Figure 2: Proportions of CD3+ T cells in the placenta and cord blood by ART group
Box plots (showing medians and interquartile ranges) of (A) CD4+ and (B) CD8+ T cells in the decidua parietalis (DPar), decidua basalis (DBas), villous tissue (VTiss) and cord blood (Cblood) from HIV-uninfected (MHIV neg) and infected mothers on efavirenz (EFV) or dolutegravir (DTG) and HIV unexposed and uninfected (HUU) and HIV exposed uninfected (HEU) cord bloods. Tests of significance were performed using the Kruskal-Wallis test.
Supplementary Figure 3: Proportions of CD4+ T cells in the placenta, cord blood and maternal viral load Correlation plots between CD4+ T cell proportions in the placenta and cord blood and maternal viral load one week after ART initiation (visit 2, 8 weeks before delivery), four weeks after ART initiation (visit 3, 4 weeks before delivery) and thirty-six weeks’ GA (visit 4, 2 weeks before delivery) in the decidua parietalis, basalis, villous tissue and cord blood. Statistical analysis was performed using the Spearman rank test and the grey shaded area represents the 95% confidence intervals.
Supplementary Figure 4: Proportions of CD8+ T cells in the placenta, cord blood and maternal viral load Correlation plots between CD8+ T cell proportions in the placenta and cord blood and maternal viral load one week after ART initiation (visit 2, 8 weeks before delivery), four weeks after ART initiation (visit 3, 4 weeks before delivery) and thirty-six weeks’ GA (visit 4, 2 weeks before delivery) in the decidua parietalis, basalis, villous tissue and cord blood. Statistical analysis was performed using the Spearman rank test and the grey shaded area represents the 95% confidence intervals.
Supplementary Figure 5: Memory differentiation of CD4+ T cells in the villous tissue and cord blood
Box plots (showing medians and interquartile ranges) of CD4+ Naïve, Early differentiated (ED), Late differentiated (LD) and Terminally differentiated (TD) T cells in the villous tissue (VT; upper panel) and cord blood (CB; lower panel) from HIV-uninfected (MHIV neg) and infected mothers (MHIV pos) and HIV unexposed and uninfected (HUU) and HIV exposed uninfected (HEU) cord bloods. Tests of significance were performed using the Mann-Whitney U test.
Supplementary Figure 6: Memory differentiation of CD4 and CD8 T cells in the decidua parietalis and decidua basalis
Representative flow cytometry contour plots of Naïve, Early differentiated (ED), Late differentiated (LD) and Terminally differentiated (TD) CD4+ T cells (upper panel) and CD8+ T cells (lower panel) in the decidua parietalis and basalis.
Supplementary Figure 7: Memory phenotype of CD4 T cells in the Villous Tissue
Box plots (showing medians and interquartile ranges) of CD4+ and CD8+ Naïve, Early differentiated (ED), Late differentiated (LD) and Terminally differentiated (TD) T cells in the decidua parietalis and basalis from HIV-uninfected (MHIV neg) and infected mothers (MHIV pos). Tests of significance were performed using the Mann-Whitney U test.
Acknowledgments
We wish to thank all the study participants in this study; all the members of the DOLPHIN-2 clinical trial and INFANT placenta study. We also wish to thank Nonzwakazi Bangani, Goitseone Thamae, Michelle Barboure, Berenice Alinde and Lizette Fick for their expert assistance in sample processing and Dr Amsha Ramburan for capturing the FISH images.
Footnotes
Conflict of interest statement The authors have declared that no conflict of interest exists.
References



