
Abstract
Visual alterations under classic psychedelics can include rich phenomenological accounts of eyes-closed imagery. Preclinical evidence suggests agonism of the 5-HT2A receptor may reduce synaptic gain to produce psychedelic-induced imagery. However, this has not been investigated in humans. To infer the directed connectivity changes to visual sensory connectivity underlying psychedelic visual imagery in healthy adults, we applied dynamic causal modelling to the resting state eyes-closed functional MRI scans of 24 subjects after administration of 0.2mg/kg of the serotonergic psychedelic drug, psilocybin (magic mushrooms), or placebo. The effective connectivity model included the early visual area, fusiform gyrus, intraparietal sulcus, and inferior frontal gyrus was modelled. We observed a pattern of increased self-inhibition of both early visual and higher visual-association regions under psilocybin that was consistent with preclinical findings. We also observed a pattern of reduced excitation and inhibition from visual-association regions to earlier visual areas that indicated top-down connectivity is enhanced during visual imagery. The results were associated with behavioural measures of complex and elementary imagery taken after the scans, suggesting psilocybin-induced deprivation of sensory activity amplifies endogenous neural activity associated with the perception of eyes-closed visual imagery. The findings inform our basic and clinical understanding of visual perception and identify neural mechanisms that contribute to psychedelic-induced eyes-closed visual imagery.
Significance statement Our study is the first measurement of the effects of a 5-HT2A receptor agonist on synaptic gain of the visual system in humans and advances the understanding of the psychopharmacology of visual perception. Our findings are consistent with preclinical evidence and elucidate the effects of psychedelics on neural mechanisms of the visual system. Specifically, we identify manifestations of visual imagery in the absence of stimuli result from decreased self-connectivity of the visual system and increased reliance on top-down connectivity. The findings have relevance to understanding clinical hallucinations and offers avenues for exploring the neurology and utility of visual imagery in psychedelic-assisted-psychotherapy. Moreover, our study advances the broader field of neuroimaging by identifying the hierarchical organisation of brain connectivity underlying perception.
Introduction
Our imagination allows us to see beyond the ordinary constraints of sensory perception. However, our capacity to visualise imagery is typically restricted during our waking state. When a serotonergic psychedelic is ingested, these restrictions can be altered. In this study, we investigate the neural mechanisms of psychedelic-induced eyes-closed visual imagery (i.e., hallucinations) using psilocybin, a prototypical classic psychedelic derived from select species of mushrooms. Psychedelics allow non-voluntary hallucinated visual imagery to be embedded in an experimental design, which may otherwise be difficult due to the unpredictable occurrence of hallucinations in clinical disorders (Teufel, 2018). This research can help us understand the relationship between neuropharmacology and large-scale neural connectivity of the visual system that informs our understanding of perception and clinical hallucinations.
Psychedelic perceptual changes are primarily derived from agonism of the serotonergic 5-HT2A receptor (5-HT2AR) (Glennon, Titeler, & McKenney, 1984; Madsen et al., 2019; K. H. Preller et al., 2017; F. X. Vollenweider, Vollenweider-Scherpenhuyzen, Babler, Vogel, & Hell, 1998). The agonism of psychedelic molecules binding to the 5-HT2AR produce an excitatory effect on layer 5 pyramidal neurons which are suggested to modulate neural gain (Burt et al 2021). The 5-HT2AR is highly expressed in the visual and frontal areas of the brain. However, psychedelics may alter neuronal activity in these areas of the brain differently. Under serotonergic psychedelics, frontal area functional connectivity — which describes the temporal synchrony of activity among brain regions—appears to desynchronise (Katrin H. Preller et al., 2020). Frontal regions serve roles in self-related thinking, planning and egocentric perspective that under psychedelics have been associated with the effect of ego dissolution (i.e., decreased self-referential awareness and perception of unity between the self and the world) (Robin L. Carhart-Harris et al., 2012; Smigielski, Scheidegger, Kometer, & Vollenweider, 2019; Franz X. Vollenweider & Preller, 2020; Stoliker et al., 2022). In contrast, visual areas of the brain undergo an opposite transformation and show increased synchrony of functional connectivity that may be associated with change to 5HT2AR (Katrin H. Preller et al., 2020). For example, evidence for the role of the 5-HT2AR in visual effects (i.e., hallucinations) has been demonstrated in Parkinson’s disease (Ballanger et al., 2010).
Psychedelic-induced visual effects can occur with eyes-open or eyes-closed. Review of eyes-closed visual effects suggests they can occur at different levels of complexity and some subjective reports suggest the visualisations can form imagery that match the vividness and realism of eyes-open stimuli (Cott & Rock, 2008; de Araujo et al., 2012; M. Kometer & Vollenweider, 2018; Shanon, 2002). The most rudimentary forms of eyes-closed visual effects include elementary imagery. Elementary imagery comprises light flashes, reorganising and moving line orientations and geometrical figures containing recurrent patterns (Oishi et al., 2003). These effects have been associated with the early visual area (EVA; corresponding to the primary visual cortex) and may be accompanied by visual intensification such as increased brightness or contrast (M. Kometer & Vollenweider, 2018; Price, Whitlock, & Hall, 1983; Teeple, Caplan, & Stern, 2009). More phenomenologically rich visual imagery can also occur under psychedelics to induce visualisations of semantic content such as scenes, people, and objects. These visualisations are identified as complex imagery and have been associated with visual associative connectivity (N. Dijkstra, Bosch, & van Gerven, 2017; Price et al., 1983; Teeple et al., 2009). Complex imagery is also a form of visual hallucination witnessed in conditions of visual impairment such as Charles Bonnet Syndrome (Manford & Andermann, 1998; Menon, Rahman, Menon, & Dutton, 2003). Complex imagery and elemental imagery under psychedelics have been assessed independently using the five dimensions of altered states of consciousness scale, which is a retrospective self-report measure commonly used in psychedelic research (Studerus, Gamma, & Vollenweider, 2010), see Supplementary for further information 1.
Preclinical research investigating change to neural mechanisms of the visual system in the mid 20th-century found that semi-synthetic psychedelic Lysergic diethylamide acid (LSD) decreased neuronal responses of the lateral geniculate nucleus in response to optical tract stimulation of anesthetised cats (Evarts, Landau, Freygang, & Marshall, 1955). These findings were supported by contemporary preclinical research that used extracellular recordings and wide-field calcium imaging, which reported imbalanced 5-HT1A/2A activation reduced sensory drive of the mouse visual cortex (Azimi et al., 2020). Similar research using wide-field two-photon calcium imaging and single-unit electrophysiology of mice under the 5-HT2AR agonist DOI (2,5-dimethoxy-4-iodoamphetamine) also identified reduced net inhibition of visual response amplitude and surround suppression (eyes-open)2, while visual feature tuning properties such as retinotopic organization and receptive fields were maintained (Michaiel, Parker, & Niell, 2019).
The influence of alpha inhibition in the visual system during eyes-closed may also be relevant to psychedelic closed-eye imagery. Basic preclinical research that measured alpha inhibition found that when our eyes are closed, alpha waves (8-12 Hz range) inhibit activity in the visual system (Billock & Tsou, 2007; Klimesch, 2011). Preclinical research in mice measured single-cell neural firing rates under activation of the 5-HT2AR and identified suppressed high firing rates that are typically indicative of visual stimuli (Watakabe et al., 2009 (M. Kometer & Vollenweider, 2018; Montemurro, Rasch, Murayama, Logothetis, & Panzeri, 2008; Quiroga, Reddy, Kreiman, Koch, & Fried, 2005). Follow-up clinical investigation of alpha inhibition under psilocybin corroborated these results by detecting inverted (i.e., dampened) response of occipital-parietal alpha oscillations to eyes-open and eyes-closed states (M. Kometer, Cahn, Andel, Carter, & Vollenweider, 2011; Michael Kometer, Schmidt, Jäncke, & Vollenweider, 2013)(Michael Kometer et al., 2013). The reduced alpha inhibition was associated with reduced attention to sensory stimuli during eyes-open and decreased alpha inhibition during eyes-closed periods. This may indicate activity in visual connectivity with eyes-closed that resembles activity typically reserved for eyes-open stimuli (Michael Kometer et al., 2013; M. Kometer & Vollenweider, 2018). Similar findings are associated with eyes-closed visual alterations under the psychedelic N,N-Dimethyltryptamine (DMT), which also identified reduced low frequencies in parietal-occipital regions associated with eyes-closed states (Timmermann et al., 2019). Increased activity in the visual system in the functional connectivity measurements of human subjects under LSD identified the early visual system responds comparable to receiving localised spatial inputs during eyes-closed (Roseman et al., 2016). These functional connectivity changes may have been induced by reduced alpha inhibition of visual connectivity during eyes-closed.
Responses of the visual system to stimulus-triggered perception and internally generated visual imagery rely on similar neural mechanisms (Fazekas, Nemeth, & Overgaard, 2020). This may suggest that increased activity of visual connectivity in eyes-closed states under psychedelics gives rise to visualisations. This idea is further reinforced by evidence that occipital-parietal regions reduce low frequencies during dreaming (Siclari et al., 2017; Timmermann et al., 2019).
To date, the available evidence suggests that psychedelics reduce stimulus-evoked responses along the primary visual pathway during eyes-open conditions. Conversely, during eyes-closed conditions, they appear to disrupt inhibitory mechanisms that control activity in the visual system and increase activity resembling the inputs of external stimuli, potentially indicating that top-down associative inputs influence visual imagery. However, the estimation of sensory inputs in the visual system under psychedelic-induced 5-HT2A receptor agonism have not been investigated in humans. Our study investigated regions of the visual system previously established to be associated with mental imagery (N. Dijkstra, Zeidman, Ondobaka, van Gerven, & Friston, 2017). The regions included the EVA, fusiform gyrus (FG), intraparietal sulcus (IPS) and inferior frontal gyrus (IFG). The EVA is the first stage of cortical visual processing and has demonstrated increased top-down connectivity from visual-association regions to the EVA during eyes-closed visual imagery tasks (N. Dijkstra, Zeidman, et al., 2017). Connectivity from the EVA follows two broad pathways. The dorsal pathway is connected with the fusiform gyrus (FG), which is part of the inferior temporal cortex that functions to recognise objects and faces (Blatt, 2013). The ventral pathway from the EVA is connected to the intraparietal sulcus (IPS), which, along with the inferior frontal gyrus (IFG), is involved in cognitive aspects of imagery. We hypothesised increased self-inhibition and reduced excitatory connectivity of these regions under psilocybin associated with eyes-closed imagery. Specifically, we expected that EVA self-connectivity would be associated with elementary imagery and top-down (i.e., feedback or inside-out) directed connectivity would be associated with complex imagery.
Results
We report effective connectivity results in the visual pathways under both placebo and psilocybin conditions and the association of the effectivity with elementary and complex imagery scores obtained after the resting state scan.
Results are reported as the mean effective connectivity of conditions without application of global signal regression (GSR). Multiple design matrices without and with GSR are reported in the supplementary material (See Fig S3, S4 and S5). In Dynamic Causal Modelling (DCM), self-connections are always modelled as inhibitory and log-scaled (see Supplementary Material for brief technical explanation). Self-connections control the regions’ gain or sensitivity to inputs and the synaptic gain or sensitivity of a region to inputs. A positive self-connection means a relative increased inhibition of the region to external inputs, whereas a negative self-connection means a relative decreased inhibition (i.e., disinhibition) and increased synaptic gain or sensitivity to inputs. Only the self-connections are log-scaled in DCM.
Visual pathway effective connectivity under placebo
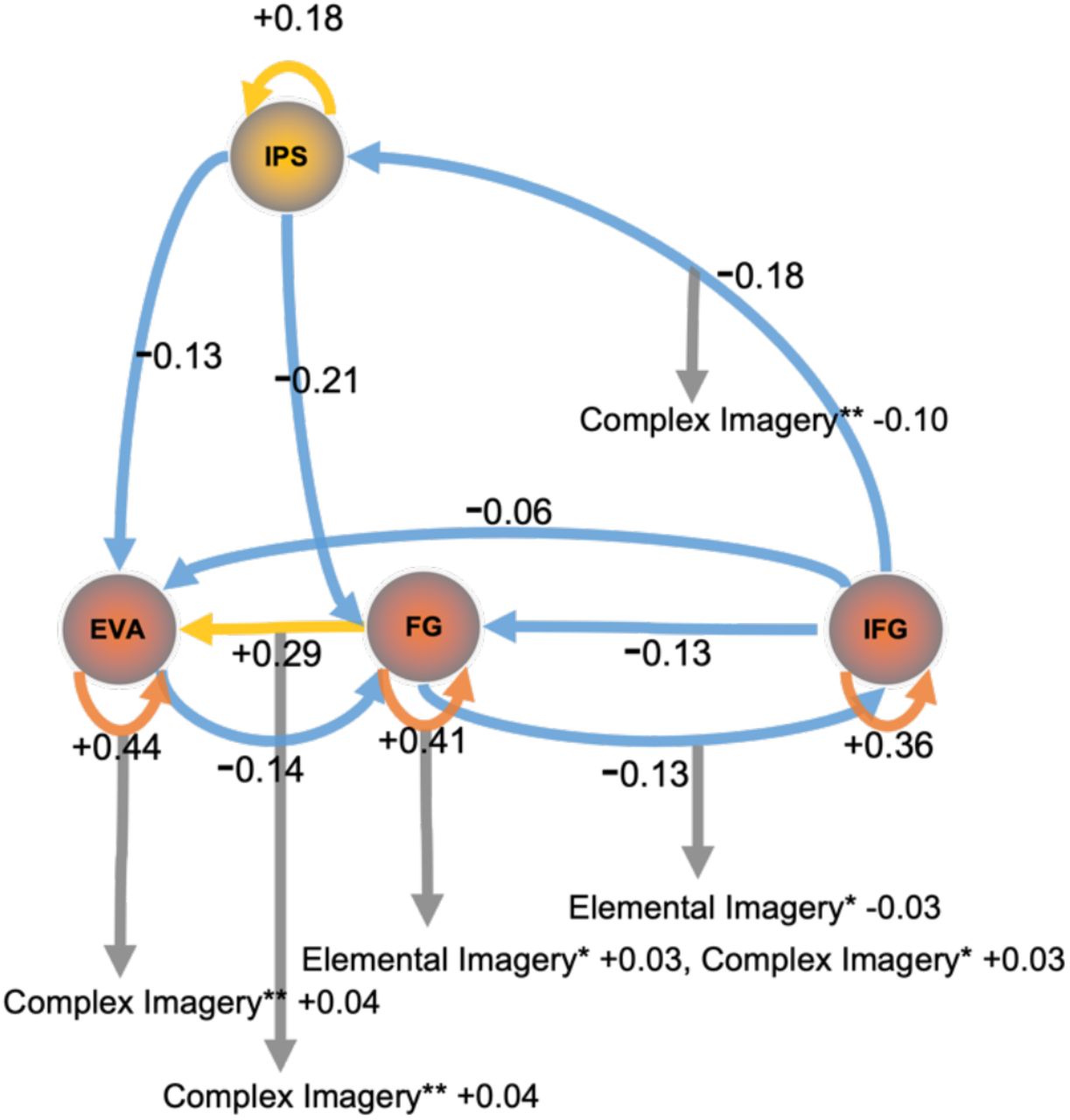
Group-level mean effective connectivity interactions between regions of the visual pathway show inhibition from the IFG to the IPS, FG and EVA. Self-inhibition of the EVA is accompanied by excitation from the FG to the EVA (Fig 1). The results were similar with and without GSR (See Fig S4 and S5 for results with global signal regression).

Mean effective connectivity 70 minutes post-administration of a placebo or psilocybin. Cool colours represent inhibition; warm colours represent excitation. Warm coloured self-connections represent increased inhibition. Left panel, mean effective connectivity 70 minutes post-administration of a placebo: Self-inhibition of the EVA and excitatory feedback connectivity to the EVA from the FG was found. Strong inhibitory connectivity was also estimated from the IFG to the IPS and FG, and to a lesser degree to the EVA. Right panel, mean effective connectivity under psilocybin 70 minutes post-administration: All region self-connections show inhibition. Inhibition is demonstrated across all connections expect the fusiform gyrus. Values display effect sizes (posterior expectations) of connections in Hz except self-connections which are modeled as always inhibitory and are log-scaled. All results are for posterior probability > 0.99 which is consistent with very strong evidence. Regions – Early Visual Area (EVA), Fusiform Gyrus (FG), Inferior Frontal Gyrus (IFG), Intraparietal Sulcus (IPS).
Visual pathways under psilocybin
Group-level mean effective connectivity between regions of the visual pathway show inhibition from the IFG to the IPS, FG and EVA, as well as between all connections to the EVA, except for the FG to EVA, which was excitatory. Self-connectivity of all regions is inhibitory, particularly within the EVA and regions of the inferior temporal cortex (FG and IFG) which showed increased inhibitory recurrent connectivity (Fig 1). See Fig S1 for detailed group level region effective connectivity matrix and Table S1 for credible intervals.
Behavioural associations to effective connectivity under psilocybin
Behavioural scores acquired after the scans measured elemental imagery and complex imagery. Associations between group-level mean effective connectivity and elemental and complex imagery were analysed. Self-inhibition of the FG and inhibitory connectivity from the FG to the IFG were associated with elemental imagery. Self-inhibition of the EVA and FG, and connectivity from the FG to EVA were associated with complex imagery. Complex imagery was also associated with the inhibition of the IFG to the IPS. See Fig 2 and Fig S2 for the respective connectivity matrices.

Behavioural associations to effective connectivity under psilocybin. Cool colours represent inhibition; warm colours represent excitation. Warm coloured self-connections represent increased inhibition. The EVA, which was self-inhibitory, was positively associated to complex imagery while the self-connectivity of the FG was positively associated to elemental and complex imagery. Excitatory feedback connectivity from the FG to EVA was also positively associated to complex imagery while inhibitory connectivity to the IFG was positively associated to elemental imagery. Elemental imagery and complex imagery scores were measured on the 5D-ASC immediately after scans and these scores were associated with effective connectivity (see supplementary). Values display effect sizes (posterior expectations) of connections in Hz except self-connections which are modeled as always inhibitory and are log-scaled. * Denotes posterior probability > 0.50, ** denotes posterior probability > 0.99 which amounts to very strong evidence.
Discussion
When we open our eyes, visual perception appears to be driven by the stimuli of the outside world. However, consciousness research has identified the significant role of the brain in constructing visual perception. The German physicist and physician, Hermann von Helmholtz said, “Objects are always imagined as being present in the field of vision as would have to be there in order to produce the same impression on the nervous mechanism”. This axiom suggests visual perception of the external world manifests from the inference of neural activity. Neural activity corresponding to vision is guided by signals from the sensory epithelia of the eyes. However, the eyes’ ability to represent the environment is limited by foveal acuity, subcortical filtration mechanisms and attention processes. Prior knowledge, recognition and context, such as visual schemas, help determine the likely causes of sensory sensations and signify the importance of beliefs and associations in the construction of visual perception. For example, inferences in visual perception rely on expectations. Expectations that influence perception may be learned, for example, prior expectations have been shown to facilitate hallucinations (Corlett et al., 2019) or prior expectations may be innate, demonstrated by visual illusions and perceptual biases, such as the Adelson’s checker-shadow illusion (Huff, Mahabadi, & Tadi, 2022). These examples and previous research demonstrate that visual perception is configured by intrinsic neural mechanisms and influenced by high-level cognitive inferences.
Our investigation measured the large-scale expression of psychedelic neuropharmacological changes on brain connectivity to give insights into the neural mechanisms that underlie the perception of eyes-closed visual imagery in human subjects under psilocybin. The findings revealed a pattern of inhibition in the effective connectivity of self-connections of visual pathways of subjects under psilocybin. DCM estimated that inhibition in the self-connections of all regions in our model under the influence of psilocybin was greater than the inhibition seen in subjects under placebo (see Figs 1 and 2). The results of the current study align with previous preclinical findings that indicate that reduced sensory drive enhances internal transmissions under agonism of the 5-HT2AR (as reported by Azimi et al., 2020; Evarts et al., 1955; Michaiel et al., 2019). The reduced sensory drive appears to generalise to the effective connectivity of the human visual system under psilocybin and suggests the augmentation of internal signals may induce visual imagery, in the absence of eyes-open sensory stimuli. The increased self-inhibition of visual and associative regions indicates a decrease in synaptic gain that corresponds to the decreased sensitivity of these regions to inputs. Additionally, it is noteworthy to mention that psilocybin reduced sensitivity to external stimuli under eyes-open conditions (Schmidt, Kometer, Bachmann, Seifritz, & Vollenweider, 2013). The decreased synaptic gain may relate to previous findings of reduced parieto-occipital alpha oscillations measured in the 8-12Hz band under psilocybin, which may facilitate spontaneous activation in eyes-closed states (as reported by Michael Kometer et al., 2013; M. Kometer & Vollenweider, 2018). Although the decreased synaptic gain appears to contradict increased activity in the visual cortex and the synchronised temporal patterns identified in sensory regions by functional connectivity analysis (as reported by Roseman et al., 2016; Katrin H. Preller et al., 2020), these discrepancies may originate from measurement techniques and analysis methods.
The self-inhibition of specific regions, such as the EVA and FG was also demonstrated in association with complex and elemental imagery. The FG is strongly involved in the processing and recognition of objects and faces (Blatt, 2013), while the EVA receives, segments, and integrates visual information (Huff et al., 2022). Visual effects related to the visual functions of EVA and FG may be associated with the self-inhibition of these regions. We also found top-down connectivity from the FG to EVA associated with complex imagery and damage to this connectivity has previously been associated with Charles Bonnet syndrome hallucinations (Pang, 2016).
Effective connections between the IFG to EVA under psilocybin showed less inhibition than placebo. Previous fMRI research of healthy adults has estimated the effective connectivity of voluntarily imagined objects (N. Dijkstra, Zeidman, et al., 2017). This research indicated group activity from the IFG to the EVA was more excitatory when objects were imagined versus when visually observed, suggesting that sensory perception is underwritten by feedforward connectivity while imagery is underwritten by feedback connectivity. The difference between our findings and that of previous research, while trending in the same direction (less inhibitory), may mark the difference between task-based imagery that requires intentional imagination and our experimental design which measured spontaneous psilocybin-induced imagery.
The IFG, which has a role in cognitive processes during visual working memory (Gazzaley & Nobre, 2012), showed greater top-down connectivity under psilocybin that is demonstrated by its efferent connections (see Fig 1). For example, we found top-down inhibitory connectivity from IFG to IPS was reduced compared to placebo and was associated with complex imagery. The IPS is involved in visual attention and the maintenance and manipulation of spatial information in working memory (Bray, Almas, Arnold, Iaria, & MacQueen, 2013). The reduced top-down inhibition under psilocybin may suggest top-down inferences are amplified and serve a role in imagery.
A popular model of psychedelic-altered consciousness, the RElaxed Beliefs Under pSychedelics (REBUS model) (R. L. Carhart-Harris & Friston, 2019; Katrin H. Preller et al., 2020), suggests reduced top-down connectivity and increased bottom-up connectivity underlies the neural mechanisms of psychedelics. While this model may apply to alternate bottom-up connectivity to the cortex (see Katrin H. Preller et al., 2019), our findings do not support the bottom-up principle of this model between visual and associative regions. Instead, we found a pattern of reduced inhibitory top-down effective connectivity and strong self-inhibition of visual and associative regions. Psilocybin-induced effects in our model are interpreted to sensitise visual connectivity to top-down endogenous signals. Greater reliance on top-down priors that we have identified is aligned with a strong priors model of the visual system that has previously been used to describe the occurrence of hallucinations (Corlett et al., 2019). Schizophrenia research which investigated different pharmacological mechanisms, also described how impaired sensory input may allow attentional mechanisms a preponderant role that leads to hallucinations (Boksa, 2009).
Therefore, our findings suggest reduced sensory drive may amplify the role of top-down inferences involved in visual perception in a manner similar to dreaming. To support this perspective we point to the reduced parieto-occipital alpha oscillations detected both in dreams and under psychedelics with eyes-closed (Michael Kometer et al., 2013); Siclari et al., 2017). Alpha waves, which are suggested to inhibit brain regions not undergoing operations, disappear during sleep, however, their role during dreams remains ambiguous (Moini & Piran, 2020; Ruby et al., 2013; Scarpelli, D’Atri, Gorgoni, Ferrara, & De Gennaro, 2015). Relatedly, the experience of sensory synaesthesias, such as visual imagery driven by music, is common under psychedelics (Kaelen et al., 2016) and suggests the delineation of signals may be diminished under psychedelics to allow top-down associative inferences to manifest to as imagery.
Signals that influence visual imagery may derive from multiple connectivities throughout the brain. For example, limbic connectivity such as between the retrosplenial cortex, anterior hippocampus, parahippocampus and entorhinal cortex that share roles in egocentric and allocentric perspectives and spatial aspects of imagery (Alexander Andrew et al.; Doeller & Kaplan, 2011; Kim & Maguire, 2018; Vann, Aggleton, & Maguire, 2009). Future connectivity research could measure interactions between visual, emotional and association connectivity to investigate the mechanisms that control the content and complexity of psychedelic imagery. This pursuit may also advance therapeutic applications using psychedelics. For example, research has described the contents of complex visual imagery can take on a personalised meaning that may support therapeutic changes (Malone et al., 2018). Psychedelics may also be useful to test with populations diagnosed with aphantasia, who ordinarily cannot perceive visual imagery.
Future research studies could measure effective connectivity changes between visual and associative regions at higher psychedelic doses to advance our understanding of the integrative function between sensory and associative connectivity. For example, it has been suggested that the difficulty to differentiate whether the source of a signal represents reality or imagination, known as reality monitoring, depends upon top-down connectivity to earlier visual areas, such as the IFG to the EVA (N. Dijkstra, Kok, P., & Fleming, S. M., 2021). Higher doses of psilocybin which result in experiences of ego dissolution and related out-of-body experiences can give insight into the mechanisms that underlie these symptoms in clinical disorders. We found the IFG, which serves a role in cognitive processes during visual working memory (Gazzaley & Nobre, 2012), was diminished under psilocybin. Connectivity between the IFG to EVA may be an important mechanism for reality monitoring that future research could investigate at increasing doses of psychedelics.
Our investigation of imagery in a healthy population under psilocybin has several limitations. The selection of brain regions and the coordinates of the regions may influence the results. Replication using the same regions is necessary. Future research may explore an alternate selection of regions and methods to determine their coordinates. For example, the cuneus, precuneus, posterior parietal, lingual, and lateral occipital gyrus are visual regions that serve important functions, often distinguished by the dorsal and ventral visual pathways, that were not investigated in this analysis. Imaging that accommodates subjective differences in the onset of visual imagery is also needed. Imagery tasks may also facilitate group-specific activations and the identification of coordinates in the regions of interest. Furthermore, the psychedelic dose can strongly impact the results. Participants were given less than a standard clinical dose of psilocybin typically used during therapeutic interventions (i.e., 25 mg). The vividness of imagery and connectivity dynamics may be more altered at a higher dose. In addition, the score distribution on our measures of the 5D-ASC may also be a limitation. Participants responded to imagery measures using a button box that constrained group scores to a 1-4 scale. The limited granularity of the score distribution may affect the analysis of behavioural associations between connectivity and imagery. However, alignment between the time of imaging and behavioural measurement ensured the altered visual experience occurred during the scan. The small healthy adult sample (n=20 after thresholding for head motion) is also a limitation. Larger group sizes may benefit the reliability of results and are recommended for future research. Lastly, preclinical research varied between anaesthetised and eyes-open recordings, further suggesting the alignment of the present results with preclinical literature requires validation from further clinical studies.
Conclusion
When we close our eyes, sensory stimulation is inhibited, and we experience a sense of separation from the outside world. In this research, we demonstrated neural mechanisms of the visual system altered by psychedelics that may cause visual perceptions in the absence of external stimulation. We found inhibition of visual brain region inputs and reduced effective connectivity effect size between regions under psilocybin associated with complex and elemental imagery. Psychedelics’ ability to inhibit and decrease the gain of sensory connectivity suggests sensory deprivation may enhance endogenous signals and underlie closed-eye imagery. Our results extend previous preclinical research that found 5-HT2AR inhibits the stimulus-evoked activity of visual regions, provides insight into clinical hallucinations, and helps describe the neural mechanisms of visual imagery. Further research with psychedelics will be required to confirm our findings using different coordinates and regions.
By altering perception, psychedelics provide a means of measuring changes in the mechanisms responsible for perceptual interpretation of the world. The ability of psychedelics to induce visual alterations and imagery expands the scope for perception and clinical research alongside the investigation of psychiatric disorders, brain injury, sleep and dreams. Imagery in closed-eye states suggests psychedelics affect the brain’s function to infer discrimination between the subjective and the objective worlds that is consistent with an overarching theme of dissolved subject-object boundaries under psychedelics.
Methods
Participants
The data analysed in this paper were collected as part of a previous study (registered at ClinicalTrials.gov (NCT03736980)), which is reported in (K. H. Preller et al., 2017) and was approved by the Cantonal Ethics Committee of Zurich. 24 subjects (12 males and 11 females; mean age = 26.30 years; range = 20–40 years) were recruited through advertisements at universities in Zurich, Switzerland. See supplementary material for further detail.
Design
The data we analyse is part of a double blind, randomised, placebo-controlled, cross-over study was performed. Resting state scans (10 minutes each) were taken 20, 40 and 70 minutes following administration of psilocybin or placebo. However, only scans at 70 minutes during the peak effects were used in our analysis. Participants were asked to not engage in repetitive thoughts such as counting. Furthermore, participants were asked to close their eyes during the resting state scan. A short version of the 5 Dimensions of Altered States of Consciousness Scale (5D-ASC, a retrospective self-report questionnaire) (Dittrich, 1998) was administered right after the scan to assess the time course of subjective effects. The questions were displayed with MR-compatible video goggles Resonance Technology Inc., Northridge, USA). Participants answered the questions on a scale ranging from 1 (not at all) to 4 (very much) using a 4-button response box.
MRI Data Acquisition and Preprocessing
MRI data were acquired on a Philips Achieva 3.0T whole-body scanner. A 32-channel receive head coil and MultiTransmit parallel radio frequency transmission was used. Images were acquired using a whole-brain gradient-echo planar imaging (EPI) sequence (repetition time, 2,430 ms; echo time, 27 ms; slice thickness, 3 mm; 45 axial slices; no slice gap; field of view, 240 × 240 mm2; in-plane resolution, 3 × 3 mm; sensitivity-encoding reduction factor, 2.0). 265 volumes were acquired per resting state scan resulting in a scan duration of 10 mins. Additionally, two high-resolution anatomical images (voxel size, 0.7 × 0.7 × 0.7 mm3) were acquired using T1-weighted and T2-weighted sequences using 3D magnetization prepared rapid-acquisition gradient echo sequence (MP-RAGE) and a turbo spin-echo sequence, respectively. See Supplementary for more details. The acquired images were analysed using SPM12 (Ashburner et al., 2014). The pre-processing steps of the images consisted of slice-timing correction, realignment, spatial normalization to the standard EPI template of the Montreal Neurological Institute (MNI), and spatial smoothing using a Gaussian kernel of 6-mm full-width at half maximum. Head motion was investigated for any excessive movement. 3 subjects were excluded due to excessive head motion and one subject did not complete the scan at 70 minutes so eventually 20 subjects entered our connectivity analysis pipeline.
Extraction of region coordinates across subjects
Regions, model complexity and ROI size were selected based on prior literature (N. Dijkstra, Zeidman, et al., 2017; Ishai, Haxby, & Ungerleider, 2002; Kalkstein, Checksfield, Jacob Bollinger, & Gazzaley, 2011; Reddy, Tsuchiya, & Serre, 2010). A generalized linear model (GLM) was used to regress 6 head motion parameters (3 translation and 3 rotational), white matter and cerebrospinal fluid signals from preprocessed data. Global signal regression (GSR) was not used in our pre-processing pipeline to align with visual research of the 2A receptor (Azimi et al., 2020; Michaiel et al., 2019). However, the results without GSR are reported in supplementary material to provide consistency with line with our previous investigations (Stoliker et al., 2022). The time series for each ROI was computed as the first principal component of the voxels activity within an 8 mm sphere centred on the region of interest (ROI) coordinates listed in Table 1.

The visual-associative regions comprised of the early visual cortex (EVC), fusiform gyrus (FG); inferior parietal sulcus (IPS) and inferior frontal gyrus (IFG).
Specification and Inversion of DCM
Dynamic causal modelling (DCM) enables measurement of the direction of connectivity among circuits, which relies on Bayesian model comparison demonstrated to discriminate between inhibition and excitation of bottom-up and top-down connections (Bastos et al., 2015; Razi, Kahan, Rees, & Friston, 2015). This analysis technique allows non-invasive inference of the directed connectivity and self-connectivity (i.e., synaptic activity).
A fully-connected DCM model was specified using the four ROIs identified in Table 1, without any exogenous inputs. The DCM for each subject was then inverted using spectral DCM (Karl J. Friston, Kahan, Biswal, & Razi, 2014; Razi et al., 2015) to infer the effective connectivity that best explains the observed cross-spectral density for each subject. The DCM fit to the data using cross spectral density averaged 87.3% for placebo conditions and 86.3% for psilocybin conditions.
Second Level Analysis Using Parametric Empirical Bayes
The effective connectivity inferred by spectral DCM for each subject is taken to the second (group) level to test hypotheses about between-subject effects. A Bayesian General Linear Model (GLM) is employed to characterise individual differences in effective connectivity into hypothesised group-average connection strengths plus any unexplained noise. Hypotheses on the group-level parameters are tested within the Parametric Empirical Bayes (PEB) framework (K. J. Friston et al., 2016), where both the expected values and the covariance of the connectivity parameters are taken into account. That is, precise parameter estimates influence the group-level result more strongly than uncertain estimates, which are down-weighted. We further employed Bayesian model reduction as an efficient automatic search to perform Bayesian model selection (K. J. Friston et al., 2016).
Data Availability
All data produced in the present study are available upon reasonable request to the authors
Footnotes
↵+ Joint Senior Authors
Revision of text. Analysis and results unchanged.
↵1 See Kometer and Vollenweider 2018 for a review of Serotonin Hallucinogen-Induced Visual Perceptual Alterations (M. Kometer & Vollenweider, 2018).
↵2 Surround suppression refers to the presence of neighbouring stimuli diminishing the neural response to a central visual stimulus in the primary visual cortex (Schallmo & Murray, 2016).




{kind=link}
{kind=link}
{kind=link}